 Un pequeño porcentaje de predadores significaría que habían tenido un buen apetito endotérmico.
Un pequeño porcentaje de predadores significaría que habían tenido un buen apetito endotérmico.En la actualidad, en algunos hábitats africanos, los predadores constituyen tan sólo entre el uno y el seis por ciento de la población animal en su totalidad.
Para los dinosaurios, a deducir de las muestras fósiles, Bakker encontró que los predadores representaban entre el uno y tres por ciento del total.
La relación presa-predador de los reptiles premesozoicos, que eran indiscutiblemente ectotérmicos, alcanzaba entre el treinta y cinco y el sesenta por ciento. Esos datos se convirtieron en la base fundamental de la campaña, cada vez más agresiva, de Bakker, en favor de la endotermia de los dinosaurios.
Presentó su defensa de la endotermia de los dinosaurios en un artículo publicado en Scientific American que tituló «Dinosaur Renaissance» («El renacimiento del dinosaurio»), en el cual afirmó: «Las relaciones predador-presa son instrumentos muy poderosos y útiles para la paleofisiología porque son los resultados directos del metabolismo de los depredadores».
Mientras tanto, Ostrom había modificado su postura en relación con la endotermia de los dinosaurios. Bakker necesitaba establecer la endotermia de los dinosaurios para reforzar su teoría que explicaba la ascendencia de los dinosaurios sobre los mamíferos en el mesozoico, suspendiendo su evolución en tanto que estos últimos tuvieron que competir con los dinosaurios. En la mente de Bakker sólo animales endotérmicos muy activos podían mantener la «superioridad» de los dinosaurios en el mesozoico.
Ostrom no había cambiado su opinión sobre el ágil Deinonychus o sobre la vitalidad de muchos otros dinosaurios; la garra, la cola y su locomoción erecta y bípeda parecían seguir siendo una prueba evidente e indiscutible en favor de su gran nivel de actividad.
Pero Ostrom no podía ir tan lejos, basándose sólo en esas pruebas, como para unirse a Bakker en sus arrasadoras generalizaciones sobre la biogenética de los dinosaurios y su superioridad. «Si Bakker se hubiese movido con mayor cautela, hubiéramos encontrado menos ardor y una mejor acogida -diría Ostrom en 1983-.
Nunca me fue posible convencerle de que la exposición mesurada siempre es superior a la exposición exagerada».
Los argumentos en favor y en contra de la endotermia se airearon en la reunión anual de la Asociación Americana para el Avance de la Ciencia, en Washington. Los partidarios de la endotermia estaban encabezados por Bakker y, en menor grado, por Ostrom y Ricqles. Nicholas Hotton III, de la Institución Smithsoniana, surgió como líder de la oposición, aunque tenía importantes aliados en Dale Russell y Philip Regal de la Universidad de Minnesota y James Spotila de la Universidad del Estado de Nueva York, en Buffalo.
Los resultados del simposio, publicados en 1980 como libro bajo el título A Coid Look at the Warm-Blooded Dinosaurs, aparecieron como un documento definitivo en la controversia.
Uno de los argumentos contra Bakker se referían al gran tamaño de los dinosaurios. Éstos se comportaban posiblemente como animales de sangre caliente no porque fuesen endotérmicos sino porque eran muy grandes. Es posible que debieran su éxito a su extraordinario tamaño, a su enorme masa.
Si hay algo en relación con los dinosaurios que está por encima de toda disputa es su tamaño. Hotton determinó que el ochenta por ciento de los mamíferos vivos son más pequeños que el más diminuto de los dinosaurios, que pesaban unos diez kilogramos, y que más de la mitad de los dinosaurios pesaban más de dos toneladas, un peso alcanzado tan sólo por el dos por ciento de los actuales mamíferos.
Parece ser, pues, que el gran tamaño fue, en cierto modo, un factor crítico para la supervivencia del dinosaurio, y los individuos mayores de una especie son los más aptos para reproducirse conduciendo a la evolución de especies de animales aún mayores, al menos hasta alcanzar cierto punto en el que las ventajas disminuyen. Algunos científicos presentaron datos sugiriendo que los ectotermos y los endotermos se hacen más parecidos entre sí a medida que aumenta su tamaño.
Los requerimientos metabólicos y consecuentemente la relación depredador-presa podrían ser muy semejantes si ambos son grandes. Según defiende Spotila, un cuerpo con una gran masa produce una temperatura corporal bastante constante.
Los dinosaurios podrían haber sido animales inactivos homeotérmicos capaces de mantener una temperatura corporal constante por cualquier medio, incluyendo aquellos dependientes del medio ambiente.
Colbert y sus colegas en sus primeros experimentos con los caimanes de Florida, habían demostrado que esos animales, descendientes de los arcosaurios, se calientan más lentamente cuando son expuestos a la luz solar y también se enfrían más lentamente que otros animales más pequeños.
Estos caimanes pueden mantener las temperaturas de sus cuerpos relativamente constantes sin necesidad de un mecanismo endotérmico o sin un aislamiento exterior, como las pieles peludas o las plumas.
Cuanto mayores son los caimanes, descubrió Colbert, más lento es el ritmo de absorción y de pérdida de calor. En el clima cálido del mesozoico las variaciones de la temperatura corporal de los animales podrían ser presumiblemente mínimas. Por consecuencia, los dinosaurios podrían tener los atributos de los animales de sangre caliente sin necesidad de ser endotérmicos.
Si hay algo en relación con los dinosaurios que está por encima de toda disputa es su tamaño. Hotton determinó que el ochenta por ciento de los mamíferos vivos son más pequeños que el más diminuto de los dinosaurios, que pesaban unos diez kilogramos, y que más de la mitad de los dinosaurios pesaban más de dos toneladas, un peso alcanzado tan sólo por el dos por ciento de los actuales mamíferos.
Parece ser, pues, que el gran tamaño fue, en cierto modo, un factor crítico para la supervivencia del dinosaurio, y los individuos mayores de una especie son los más aptos para reproducirse conduciendo a la evolución de especies de animales aún mayores, al menos hasta alcanzar cierto punto en el que las ventajas disminuyen. Algunos científicos presentaron datos sugiriendo que los ectotermos y los endotermos se hacen más parecidos entre sí a medida que aumenta su tamaño.
Los requerimientos metabólicos y consecuentemente la relación depredador-presa podrían ser muy semejantes si ambos son grandes. Según defiende Spotila, un cuerpo con una gran masa produce una temperatura corporal bastante constante.
Los dinosaurios podrían haber sido animales inactivos homeotérmicos capaces de mantener una temperatura corporal constante por cualquier medio, incluyendo aquellos dependientes del medio ambiente.
Colbert y sus colegas en sus primeros experimentos con los caimanes de Florida, habían demostrado que esos animales, descendientes de los arcosaurios, se calientan más lentamente cuando son expuestos a la luz solar y también se enfrían más lentamente que otros animales más pequeños.
Estos caimanes pueden mantener las temperaturas de sus cuerpos relativamente constantes sin necesidad de un mecanismo endotérmico o sin un aislamiento exterior, como las pieles peludas o las plumas.
Cuanto mayores son los caimanes, descubrió Colbert, más lento es el ritmo de absorción y de pérdida de calor. En el clima cálido del mesozoico las variaciones de la temperatura corporal de los animales podrían ser presumiblemente mínimas. Por consecuencia, los dinosaurios podrían tener los atributos de los animales de sangre caliente sin necesidad de ser endotérmicos.
Hotton cree que fue la homeotermia del dinosaurio la clave: «Los dinosaurios, al igual que los mamíferos, aumentan su capacidad de actividad continuada reduciendo su dependencia del medio ambiente físico como fuente de calor corporal. Ello lo hicieron así a bajo coste, mediante un sistema de conservación de calor que configura un estilo de vida que fue muy diferente de los estilos de vida de otros animales, condicionados por el sistema de generación de calor de los mamíferos que requiere un alto coste.
Los mecanismos locomotores y el tamaño ilustran una diferencia fundamental: la actividad de los dinosaurios era más tranquila que la de los mamíferos. En general la estrategia básica de los dinosaurios era “lenta y utilitaria” y lo que perdía en impulso en relación con los mamíferos lo ganaba en economía». Una parte de la estrategia, sugirió Hotton, involucra probablemente migraciones estacionales sobre distancias que sobrepasaban los 3.200 kilómetros.
En Norteamérica se han encontrado fósiles de dinosaurios en latitudes tan nórdicas como el Territorio del Yukón, a sesenta grados de latitud norte. Incluso en el suave clima del mesozoico, el Ártico no era probablemente el lugar más adecuado para los dinosaurios, sobre todo en invierno, debido al frío y a la oscuridad que interrumpían el crecimiento de las plantas.
Los grandes dinosaurios, tanto herbívoros como carnívoros, tuvieron que emigrar para poder conseguir suficiente alimento para mantenerse en acción, incluso viviendo al modesto ritmo del metabolismo ectotérmico. Hotton ofreció otras dos razones más para explicar esas migraciones.
La primera era que la propia actividad migratoria podría facilitar una fuente de calor interno en la que se podía confiar; la segunda, que los viajes los habrían mantenido expuestos aproximadamente a la misma temperatura durante todo el año, lo que significaba una consideración vital si, en su calidad de animales ectotérmicos, la tolerancia de los dinosaurios a las fluctuaciones térmicas estaba limitada dentro de unos márgenes relativamente pequeños de emisión de calor de un metabolismo en descanso.
El concepto de que algunos dinosaurios, como muchas aves que aún viven en la actualidad, emigraran al norte o al sur, estación tras estación, es algo sorprendente pero no imposible de creer. Hotton también participaba de la opinión de Bakker de que la endotermia podía haber sido un factor que, conjuntamente con el gran tamaño y la piel desnuda de los dinosaurios, participara en las causas de su extinción. Bakker había afirmado:
«Frente a un estrés causado por un frío repentino y prolongado, los dinosaurios eran demasiado voluminosos para escapar hibernando en madrigueras o en otros microhabitáculos que estaban a disposición de los demás animales endotérmicos, y, por otra parte, eran incapaces de sobrevivir a prolongados descensos de su temperatura corporal, como ocurre con algunos tipos de tortugas, lagártidos y otros ectotermos.
La exposición a un frío severo y prolongado probablemente acabaría con la mayor parte de los endotermos vivientes, desnudos y tropicales, como los rinos, hipos, elefantes y armadillos, y también es muy posible que eliminara a los dinosaurios, los desnudos endotermos tropicales del mesozoico».
Los mecanismos locomotores y el tamaño ilustran una diferencia fundamental: la actividad de los dinosaurios era más tranquila que la de los mamíferos. En general la estrategia básica de los dinosaurios era “lenta y utilitaria” y lo que perdía en impulso en relación con los mamíferos lo ganaba en economía». Una parte de la estrategia, sugirió Hotton, involucra probablemente migraciones estacionales sobre distancias que sobrepasaban los 3.200 kilómetros.
En Norteamérica se han encontrado fósiles de dinosaurios en latitudes tan nórdicas como el Territorio del Yukón, a sesenta grados de latitud norte. Incluso en el suave clima del mesozoico, el Ártico no era probablemente el lugar más adecuado para los dinosaurios, sobre todo en invierno, debido al frío y a la oscuridad que interrumpían el crecimiento de las plantas.
Los grandes dinosaurios, tanto herbívoros como carnívoros, tuvieron que emigrar para poder conseguir suficiente alimento para mantenerse en acción, incluso viviendo al modesto ritmo del metabolismo ectotérmico. Hotton ofreció otras dos razones más para explicar esas migraciones.
La primera era que la propia actividad migratoria podría facilitar una fuente de calor interno en la que se podía confiar; la segunda, que los viajes los habrían mantenido expuestos aproximadamente a la misma temperatura durante todo el año, lo que significaba una consideración vital si, en su calidad de animales ectotérmicos, la tolerancia de los dinosaurios a las fluctuaciones térmicas estaba limitada dentro de unos márgenes relativamente pequeños de emisión de calor de un metabolismo en descanso.
El concepto de que algunos dinosaurios, como muchas aves que aún viven en la actualidad, emigraran al norte o al sur, estación tras estación, es algo sorprendente pero no imposible de creer. Hotton también participaba de la opinión de Bakker de que la endotermia podía haber sido un factor que, conjuntamente con el gran tamaño y la piel desnuda de los dinosaurios, participara en las causas de su extinción. Bakker había afirmado:
«Frente a un estrés causado por un frío repentino y prolongado, los dinosaurios eran demasiado voluminosos para escapar hibernando en madrigueras o en otros microhabitáculos que estaban a disposición de los demás animales endotérmicos, y, por otra parte, eran incapaces de sobrevivir a prolongados descensos de su temperatura corporal, como ocurre con algunos tipos de tortugas, lagártidos y otros ectotermos.
La exposición a un frío severo y prolongado probablemente acabaría con la mayor parte de los endotermos vivientes, desnudos y tropicales, como los rinos, hipos, elefantes y armadillos, y también es muy posible que eliminara a los dinosaurios, los desnudos endotermos tropicales del mesozoico».
Por el contrario, dice Hotton, «si los dinosaurios hubiesen sido endotérmicos, algunos de ellos habrían sobrevivido».
Si algunos de los pequeños dinosaurios hubieran dispuesto de plumas como aislante contra el frío, sobre lo cual se ha especulado mucho, pero nunca ha sido definitivamente establecido, habrían sobrevivido «en una diversidad comparable a la de los mamíferos y las aves supervivientes».
Incluso algunos especímenes de los dinosaurios grandes y desnudos hubieran sobrevivido, aunque sólo fuera durante un corto tiempo. Sin embargo ningún dinosaurio supuestamente sobrevivió a la extinción masiva que tuvo lugar a fines del período cretáceo. Hotton concluye:
«La supervivencia de los ectotermos, al igual que la de los endotermos, durante la transición cretáceo-terciaria, refuerza aún más el punto de vista de que existía una gran diferencia en la fisiología termal entre los dinosaurios y los tetrápodos vivientes».
Aunque los argumentos en favor del gran tamaño y la homeotermia inercial como características controladoras de la fisiología del dinosaurio, parecen satisfacer a muchos paleontólogos, Bakker no emprendió la retirada.
Los dinosaurios más pequeños se contaban entre los más ágiles y vitales de la especie, como ocurre en los endotermos. El primer grupo de tetrápodos que daba pruebas de elevada producción de calor, los terápsidos, pronto dieron auge a los grandes animales terrestres dominantes. Los dinosaurios parecen representar un elevado nivel de endotermia y reemplazaron a las dinastías de terápsidostecodontos.
La entera historia de los fósiles de los grandes vertebrados muestra la naturaleza progresiva de la evolución entre los grandes tetrápodos. Los dinosaurios parecen haber alcanzado un nivel de rendimiento en la regulación térmica y en la locomoción comparable a la de muchos de los mamíferos del cenozoico posterior.
Algunos de los pequeños dinosaurios depredadores parecen haber llegado a desarrollar cerebros del tamaño y la complejidad semejantes a los de las modernas aves no voladoras del mismo peso. Así reinterpretado, el éxito de los dinosaurios se convierte en parte de una progresión irreversible que conduce desde los primeros tetrápodos terrestres del devónico a la gran complejidad de los mamíferos modernos.
Pareció establecerse el consenso en que no existía una simple estrategia termorreguladora común a todos los dinosaurios. Los grandes dinosaurios, los saurópodos, como el Brontosaurus, probablemente estaban más cerca de la ectotermia. Sus huesos sugerían que no fueron animales muy activos o ágiles y resulta difícil imaginar cómo los saurópodos podrían haber encontrado alimento suficiente para mantener un metabolismo elevado.
La mayor parte de los dinosaurios, debido a su gran tamaño y al clima suave, probablemente mantuvieron temperaturas corporales bastante constantes. Eran homeotérmicos inerciales. Los pequeños dinosaurios, cuyos huesos sugieren una buena capacidad para la velocidad y la agilidad, pudieron haber sido endotérmicos reales. Desde luego los más pequeños como el Compsognathus, que fue un contemporáneo delArchaeopteryx, eran demasiado pequeños para poder ser homeotérmicos inerciales.
Si algunos de los pequeños dinosaurios hubieran dispuesto de plumas como aislante contra el frío, sobre lo cual se ha especulado mucho, pero nunca ha sido definitivamente establecido, habrían sobrevivido «en una diversidad comparable a la de los mamíferos y las aves supervivientes».
Incluso algunos especímenes de los dinosaurios grandes y desnudos hubieran sobrevivido, aunque sólo fuera durante un corto tiempo. Sin embargo ningún dinosaurio supuestamente sobrevivió a la extinción masiva que tuvo lugar a fines del período cretáceo. Hotton concluye:
«La supervivencia de los ectotermos, al igual que la de los endotermos, durante la transición cretáceo-terciaria, refuerza aún más el punto de vista de que existía una gran diferencia en la fisiología termal entre los dinosaurios y los tetrápodos vivientes».
Aunque los argumentos en favor del gran tamaño y la homeotermia inercial como características controladoras de la fisiología del dinosaurio, parecen satisfacer a muchos paleontólogos, Bakker no emprendió la retirada.
Los dinosaurios más pequeños se contaban entre los más ágiles y vitales de la especie, como ocurre en los endotermos. El primer grupo de tetrápodos que daba pruebas de elevada producción de calor, los terápsidos, pronto dieron auge a los grandes animales terrestres dominantes. Los dinosaurios parecen representar un elevado nivel de endotermia y reemplazaron a las dinastías de terápsidostecodontos.
La entera historia de los fósiles de los grandes vertebrados muestra la naturaleza progresiva de la evolución entre los grandes tetrápodos. Los dinosaurios parecen haber alcanzado un nivel de rendimiento en la regulación térmica y en la locomoción comparable a la de muchos de los mamíferos del cenozoico posterior.
Algunos de los pequeños dinosaurios depredadores parecen haber llegado a desarrollar cerebros del tamaño y la complejidad semejantes a los de las modernas aves no voladoras del mismo peso. Así reinterpretado, el éxito de los dinosaurios se convierte en parte de una progresión irreversible que conduce desde los primeros tetrápodos terrestres del devónico a la gran complejidad de los mamíferos modernos.
Pareció establecerse el consenso en que no existía una simple estrategia termorreguladora común a todos los dinosaurios. Los grandes dinosaurios, los saurópodos, como el Brontosaurus, probablemente estaban más cerca de la ectotermia. Sus huesos sugerían que no fueron animales muy activos o ágiles y resulta difícil imaginar cómo los saurópodos podrían haber encontrado alimento suficiente para mantener un metabolismo elevado.
La mayor parte de los dinosaurios, debido a su gran tamaño y al clima suave, probablemente mantuvieron temperaturas corporales bastante constantes. Eran homeotérmicos inerciales. Los pequeños dinosaurios, cuyos huesos sugieren una buena capacidad para la velocidad y la agilidad, pudieron haber sido endotérmicos reales. Desde luego los más pequeños como el Compsognathus, que fue un contemporáneo delArchaeopteryx, eran demasiado pequeños para poder ser homeotérmicos inerciales.
La maravilla de los dinosaurios es que son un enigma que parece por encima de toda solución. La ciencia ha identificado cientos de especies de dinosaurios, ha ensamblado sus huesos y encontrado la fecha en el tiempo de su presencia en la Tierra, pero son muchas las cosas relacionadas con las vidas de esas criaturas extrañas y monstruosas que desafían toda explicación. Los científicos salieron del debate sobre la sangre caliente al menos con un nuevo concepto del enigma del dinosaurio.
Bakker, aunque fracasó en su empeño de probar que los dinosaurios frieron endotérmicos, inspiró a un abundante grupo de paleontólogos a la exploración de la ecología y la estructura comunitaria de esos reptiles tan poco corrientes y a tratar de comprender su lugar en la evolución de la vida.
Incluso Nicholas Hotton, el más duro de los adversarios de Bakker en aquellos debates, tuvo que conceder: «Las estrategias termales alternativas y el estilo de vida al alcance de los dinosaurios pueden muy bien haber sido tan exóticos como las formas de sus cuerpos, de las que no hemos visto nunca nada comparable».
Ostrom creía firmemente que las aves son descendientes directos de los dinosaurios. Su estudio del Deinonychus lo llevó a un análisis de docenas de otros dinosaurios, en particular de los pequeños carnívoros corredores conocidos como terópodos coelurosaurios. Las similitudes existentes entre el Archaeopteryx y los coelurosaurios fueron a su juicio sorprendentes. Con la excepción de las plumas el Archaeopteryx se parece más a un pequeño dinosaurio que a un ave moderna.
Por lo tanto, concluyó Ostrom, debía haber una estrecha relación entre dinosaurios y aves, quizá más de lo que nadie se había atrevido a imaginar. Ostrom estaba resucitando la propuesta original de Thomas Henry Huxley de que existía un lazo entre los dinosaurios y las aves. Los científicos pudieron llegar a un consenso: las aves descendían de los reptiles.
Durante muchos años del siglo XX, se abandonó la idea de Huxley de un ascendiente común. Los dinosaurios y las aves estaban remotamente emparentadas debido a que ambas especies descendían de un tronco común muy distante, identificado usualmente como los primitivos tecodontos. Éstos serían los ascendientes que a principios del triásico dieron lugar a todos los arcosaurios.
Las aves, pues, no estaban más relacionadas con los dinosaurios que los cocodrilos lo están con los mismos dinosaurios o con los pterosaurios. En el caso de evolución paralela, dos organismos surgidos de un ascendiente común, divergen en dos ramas distintas que desarrollan semejanzas estructurales para adquirir capacidades comunes, como por ejemplo el vuelo.
Las aves y los pterosaurios son ejemplos de evolución paralela. En el caso de evolución convergente, los animales de dos ramas independientes y sin ningún antepasado común, tales como murciélagos y aves, desarrollan similitudes al adaptarse al mismo tipo de nicho ecológicos, como el aire.
Dado que algunos de los pequeños dinosaurios carnívoros y las protoaves compartían nichos ecológicos semejantes y perseguían el mismo tipo de alimento, es comprensible que desarrollaran anatomías similares. Pero de acuerdo con esa hipótesis eso no debía ser tomado necesariamente como signo de próximo parentesco.
Bakker, aunque fracasó en su empeño de probar que los dinosaurios frieron endotérmicos, inspiró a un abundante grupo de paleontólogos a la exploración de la ecología y la estructura comunitaria de esos reptiles tan poco corrientes y a tratar de comprender su lugar en la evolución de la vida.
Incluso Nicholas Hotton, el más duro de los adversarios de Bakker en aquellos debates, tuvo que conceder: «Las estrategias termales alternativas y el estilo de vida al alcance de los dinosaurios pueden muy bien haber sido tan exóticos como las formas de sus cuerpos, de las que no hemos visto nunca nada comparable».
Ostrom creía firmemente que las aves son descendientes directos de los dinosaurios. Su estudio del Deinonychus lo llevó a un análisis de docenas de otros dinosaurios, en particular de los pequeños carnívoros corredores conocidos como terópodos coelurosaurios. Las similitudes existentes entre el Archaeopteryx y los coelurosaurios fueron a su juicio sorprendentes. Con la excepción de las plumas el Archaeopteryx se parece más a un pequeño dinosaurio que a un ave moderna.
Por lo tanto, concluyó Ostrom, debía haber una estrecha relación entre dinosaurios y aves, quizá más de lo que nadie se había atrevido a imaginar. Ostrom estaba resucitando la propuesta original de Thomas Henry Huxley de que existía un lazo entre los dinosaurios y las aves. Los científicos pudieron llegar a un consenso: las aves descendían de los reptiles.
Durante muchos años del siglo XX, se abandonó la idea de Huxley de un ascendiente común. Los dinosaurios y las aves estaban remotamente emparentadas debido a que ambas especies descendían de un tronco común muy distante, identificado usualmente como los primitivos tecodontos. Éstos serían los ascendientes que a principios del triásico dieron lugar a todos los arcosaurios.
Las aves, pues, no estaban más relacionadas con los dinosaurios que los cocodrilos lo están con los mismos dinosaurios o con los pterosaurios. En el caso de evolución paralela, dos organismos surgidos de un ascendiente común, divergen en dos ramas distintas que desarrollan semejanzas estructurales para adquirir capacidades comunes, como por ejemplo el vuelo.
Las aves y los pterosaurios son ejemplos de evolución paralela. En el caso de evolución convergente, los animales de dos ramas independientes y sin ningún antepasado común, tales como murciélagos y aves, desarrollan similitudes al adaptarse al mismo tipo de nicho ecológicos, como el aire.
Dado que algunos de los pequeños dinosaurios carnívoros y las protoaves compartían nichos ecológicos semejantes y perseguían el mismo tipo de alimento, es comprensible que desarrollaran anatomías similares. Pero de acuerdo con esa hipótesis eso no debía ser tomado necesariamente como signo de próximo parentesco.
Gerhard Heilmann, un paleontólogo danés, fue responsable de la extendida negativa a aceptar la existencia de un lazo directo entre el dinosaurio y el ave. En su libro más importante, The Origin of Birds, publicado en 1926, Heilmann reconoció «los sorprendentes puntos de semejanza» entre los coelurosaurios y las aves.
Destacó uno de los muchos rasgos semejantes a los de las aves que se daban en el coelurosaurio: «Huesos huecos de estructura muy ligera, miembros traseros excesivamente largos con metatarsales fuertemente prolongados y un “dedo trasero”, una mano larga y estrecha, una cola larga y un cuello largo, órbitas largas y costillas ventrales». Pero faltaba uno de los rasgos más característicos de las aves.
Observó que el coelurosaurio estaba falto de toda evidencia de los huesos del cuello (clavículas). Si elcoelurosaurio no tenía clavículas ¿cómo podían sus supuestos descendientes avícolas haberlo desarrollado?
La ausencia de clavículas en el coelurosaurio, decidió Heilmann, «sería en sí misma prueba suficiente de que esos saurios no pueden ser los antepasados de las aves». En su lugar sugirió que los antepasados de las aves podrían encontrarse entre los pseudosuchianos, un grupo de tecodontos de principios del triásico.
Ya antes, en 1913, Robert Broom había encontrado en África del Sur un pequeño pseudosuchiano bípedo que tenía una antigüedad de doscientos treinta millones de años, el Euparkeria, que parecía contar con todas las cualificaciones anatómicas necesarias para ser el antepasado de las aves. Primero en 1973 y más tarde en 1975, Ostrom se incorporó al debate con firmes argumentos en favor de la existencia de un lazo directo entre los dinosaurios y las aves.
En parte lo hizo en respuesta a otra propuesta de un antepasado para las aves que negaba todo parentesco con los dinosaurios. Alick D. Walker, de la Universidad de Newcastle, sugirió que cocodrilos y aves, incluyendo elArchaeopteryx, procedían de un antepasado común teocodonto, pero que no era el mismo que daría lugar a los dinosaurios.
Basaba esta idea en el parecido entre la caja cerebral y el cráneo de los cocodrilos actuales y las aves, así como algunos notables rasgos propios de las aves que se daban en el Sphenosuchus, una criatura de finales del triásico, hace doscientos veinte millones de años, que se pensaba era un cocodrilo primitivo o un tecodonto en el camino evolutivo hacia su transformación en cocodrilo.
Ostrom pudo mostrarse más concluyenter de lo que lo había sido Huxley debido a que para entonces ya se habían descubierto y examinado muchos más dinosaurios pequeños, bípedos. Casi invariablemente sus extremidades, pies y garras, eran casi iguales a las del Archaeopteryx. U
nos cuantos de loscoelurosaurios recién descubiertos parecían poseer el desaparecido ingrediente que tanto pareció importarle a Heilmann y, por lo visto, tenían clavículas.
Una expedición polaco-mongol acababa de regresar con la prueba de que el Velociraptor, encontrado en el desierto de Gobi, parecía tener un par de huesos del cuello o clavículas. Lo mismo ocurría con el Segisaurus, un fósil norteamericano estudiado por Ostrom.
Destacó uno de los muchos rasgos semejantes a los de las aves que se daban en el coelurosaurio: «Huesos huecos de estructura muy ligera, miembros traseros excesivamente largos con metatarsales fuertemente prolongados y un “dedo trasero”, una mano larga y estrecha, una cola larga y un cuello largo, órbitas largas y costillas ventrales». Pero faltaba uno de los rasgos más característicos de las aves.
Observó que el coelurosaurio estaba falto de toda evidencia de los huesos del cuello (clavículas). Si elcoelurosaurio no tenía clavículas ¿cómo podían sus supuestos descendientes avícolas haberlo desarrollado?
La ausencia de clavículas en el coelurosaurio, decidió Heilmann, «sería en sí misma prueba suficiente de que esos saurios no pueden ser los antepasados de las aves». En su lugar sugirió que los antepasados de las aves podrían encontrarse entre los pseudosuchianos, un grupo de tecodontos de principios del triásico.
Ya antes, en 1913, Robert Broom había encontrado en África del Sur un pequeño pseudosuchiano bípedo que tenía una antigüedad de doscientos treinta millones de años, el Euparkeria, que parecía contar con todas las cualificaciones anatómicas necesarias para ser el antepasado de las aves. Primero en 1973 y más tarde en 1975, Ostrom se incorporó al debate con firmes argumentos en favor de la existencia de un lazo directo entre los dinosaurios y las aves.
En parte lo hizo en respuesta a otra propuesta de un antepasado para las aves que negaba todo parentesco con los dinosaurios. Alick D. Walker, de la Universidad de Newcastle, sugirió que cocodrilos y aves, incluyendo elArchaeopteryx, procedían de un antepasado común teocodonto, pero que no era el mismo que daría lugar a los dinosaurios.
Basaba esta idea en el parecido entre la caja cerebral y el cráneo de los cocodrilos actuales y las aves, así como algunos notables rasgos propios de las aves que se daban en el Sphenosuchus, una criatura de finales del triásico, hace doscientos veinte millones de años, que se pensaba era un cocodrilo primitivo o un tecodonto en el camino evolutivo hacia su transformación en cocodrilo.
Ostrom pudo mostrarse más concluyenter de lo que lo había sido Huxley debido a que para entonces ya se habían descubierto y examinado muchos más dinosaurios pequeños, bípedos. Casi invariablemente sus extremidades, pies y garras, eran casi iguales a las del Archaeopteryx. U
nos cuantos de loscoelurosaurios recién descubiertos parecían poseer el desaparecido ingrediente que tanto pareció importarle a Heilmann y, por lo visto, tenían clavículas.
Una expedición polaco-mongol acababa de regresar con la prueba de que el Velociraptor, encontrado en el desierto de Gobi, parecía tener un par de huesos del cuello o clavículas. Lo mismo ocurría con el Segisaurus, un fósil norteamericano estudiado por Ostrom.

«Es más probable que el Archaeopteryx adquiriera su gran número de caracteres derivados de los terópodos, por convergencia o en paralelo al mismo tiempo que esos mismos caracteres estaban siendo adquiridos por algunos terópodos coelurosaurios, presumiblemente de un común antepasado.
¿O era más factible que esos muchos rasgos derivados fueran comunes a algunos terópodos pequeños y al Archaeopteryx debido a que este último procedía directamente de un terópodo semejante? No existe en absoluto ningún reparo en mi mente para aceptar que la última de las explicaciones es, con mucho, la más probable».
La nueva actitud de Ostrom sobre el Archaeopteryx provocó un nuevo asalto en el debate de otra cuestión que hacía tiempo que dividía a los científicos. Se refería a uno de los mayores misterios de la evolución: el origen del vuelo en los animales. Como adaptación, el vuelo se coloca al mismo nivel que la salida del pez del mar, hace más de trescientos cincuenta millones de años, para convertirse en el primero de los vertebrados terrestres.
El registro de los fósiles nos muestra que el primer vuelo real se desarrolló independientemente en varias ocasiones. Los primeros animales voladores fueron los insectos alados, que aparecieron hace unos trescientos cincuenta millones de años. Después fueron los pterosaurios que, probablemente, eran capaces de efectuar un vuelo motriz.
Después llegaron las aves voladoras, quizá en una época tan antigua como la del Archaeopteryx, pero ciertamente no mucho después en el mesozoico. Los únicos otros animales voladores de propulsión propia, los murciélagos, cruzaron el aire por vez primera hace sólo cincuenta millones de años.
Aquellos que se han sentido interesados y sorprendidos por el origen del vuelo han fijado su atención en las aves, las más llamativas y logradas de todas las criaturas aéreas. El descubrimiento del Archaeopteryx facilitó a los científicos el más claro y reconocido ejemplo de una criatura próxima o en plena transición en relación a la capacidad de vuelo.
En 1880, Marsh sugirió que algunos reptiles treparon a los árboles para situar en ellos sus nidos, para alimentarse y para escapar de los depredadores. Esos primitivos antepasados de las aves tenían necesidad de saltar de una rama a otra y después de un árbol a otro. Consecuentemente cualquier variación anatómica que aumentara ese tipo de movilidad pasó a las subsiguientes generaciones por las leyes de la selección natural.
De ese modo, presumiblemente los reptiles desarrollaron plumas rudimentarias en sus antebrazos para descender cuando saltaban de rama en rama y para caer al suelo como con la ayuda de un paracaídas.
Al señalar ese punto Marsh citó el modelo de los modernos animales planeadores, como por ejemplo la llamada ardilla voladora. Más tarde, de acuerdo con la hipótesis de Marsh, estos reptiles protoavícolas, pudieron haber adaptado esas alas y fortalecido sus músculos para conseguir un vuelo impulsado muscularmente.
Otros investigadores consideran que durante el gran espacio de tiempo abarcado por el triásico, del cual apenas si tenemos registros eficaces, debió de producirse un alargamiento gradual de los dedos externos y un gran desarrollo de las capacidades trepadoras, con lo cual se ayudaba al animal a correr más. El siguiente cambio hacia las plumas pudo haber sido sencillo. Las alas debieron de ser, al principio, utilizadas para correr, después para saltar y descender desde las alturas y, finalmente, para remontarse.
Franz von Nopcsa argumentó que las aves descendían de criaturas bípedas semejantes a los dinosaurios, que corrían en el suelo. Creía que para incrementar su velocidad de carrera aquellas protoaves desarrollaron extremidades anteriores con plumas, que podían mover hacia adelante y hacia atrás como remos.
Probablemente esa habilidad no sólo permitió a aquellos animales correr con mayor rapidez sino que les dio la fuerza suficiente para elevarse y planear sobre distancias cortas. Nopcsa imaginaba que éste pudo ser el origen del vuelo de las aves. Pero eso resultaba aerodinámicamente imposible, como señaló un crítico.
Tan pronto como el animal despegara del suelo sus patas no podrían seguir propulsándolo hacia adelante y caería de inmediato. Los científicos parecían sentirse más cómodos con la teoría arbórea. Podían imaginarse con mayor facilidad que los animales hubieran tratado de utilizar la gravedad para descender de las ramas que verlos luchando contra esta fuerza para elevarse del suelo.
Tuvo que resultar importante, razonaron, el hecho de que todos los actuales vertebrados que vuelan son arbóreos. En sus estudios sobre el Archaeopteryx, Ostrom fue más allá de limitarse a estudiar su parentesco con los dinosaurios para presentar la cuestión del vuelo de las aves.
Puesto que el Archaeopteryx carecía de esternón, el hueso pectoral que liga los músculos del vuelo, sintió dudas razonables sobre su capacidad, como ave primitiva, para desarrollar un vuelo monitorizado. Creía, más bien, que se trataba de un animal «ligero de pies, rápido y que probablemente habitaba en tierra y no en los árboles, como ocurre actualmente con los faisanes, las perdices o las codornices, por ejemplo, con cuyas garras las suyas tenían un extraordinario parecido».
Todo eso le sugirió a Ostrom una nueva idea y especuló con que elArchaeopteryx y otras protoaves hubiesen sido depredadores que cazaban levantando sus presas, generalmente insectos, golpeándolas en el aire al cortarles el camino. Sus antebrazos emplumados, propuso, debían ser utilizados para golpear a los insectos en vuelo.
Cuando la superficie emplumada de sus alas se desarrolló hasta alcanzar un mayor tamaño, esto les pudo conferir la capacidad de elevarse en el aire durante sus actividades depredadoras y, con el transcurso del tiempo, hizo posible que aquellas criaturas evolucionaran en el aire con vuelo a impulso propio.
Gerald Caple, un farmacéutico, encontró poco probable que la red para cazar insectos pudiera haber evolucionado hasta dar impulso elevatorio a las capacidades necesarias para el vuelo activo motor, mostraron cómo los corredores bípedos podían desarrollar la fuerza y la capacidad suficiente para volar.
Afirmaron que tales animales, corriendo y abalanzándose sobre sus presas, podrían haber utilizado sus extremidades anteriores para guardar el equilibrio, como hacen los seres humanos cuando dan un gran salto de longitud.
Probablemente esa habilidad no sólo permitió a aquellos animales correr con mayor rapidez sino que les dio la fuerza suficiente para elevarse y planear sobre distancias cortas. Nopcsa imaginaba que éste pudo ser el origen del vuelo de las aves. Pero eso resultaba aerodinámicamente imposible, como señaló un crítico.
Tan pronto como el animal despegara del suelo sus patas no podrían seguir propulsándolo hacia adelante y caería de inmediato. Los científicos parecían sentirse más cómodos con la teoría arbórea. Podían imaginarse con mayor facilidad que los animales hubieran tratado de utilizar la gravedad para descender de las ramas que verlos luchando contra esta fuerza para elevarse del suelo.
Tuvo que resultar importante, razonaron, el hecho de que todos los actuales vertebrados que vuelan son arbóreos. En sus estudios sobre el Archaeopteryx, Ostrom fue más allá de limitarse a estudiar su parentesco con los dinosaurios para presentar la cuestión del vuelo de las aves.
Puesto que el Archaeopteryx carecía de esternón, el hueso pectoral que liga los músculos del vuelo, sintió dudas razonables sobre su capacidad, como ave primitiva, para desarrollar un vuelo monitorizado. Creía, más bien, que se trataba de un animal «ligero de pies, rápido y que probablemente habitaba en tierra y no en los árboles, como ocurre actualmente con los faisanes, las perdices o las codornices, por ejemplo, con cuyas garras las suyas tenían un extraordinario parecido».
Todo eso le sugirió a Ostrom una nueva idea y especuló con que elArchaeopteryx y otras protoaves hubiesen sido depredadores que cazaban levantando sus presas, generalmente insectos, golpeándolas en el aire al cortarles el camino. Sus antebrazos emplumados, propuso, debían ser utilizados para golpear a los insectos en vuelo.
Cuando la superficie emplumada de sus alas se desarrolló hasta alcanzar un mayor tamaño, esto les pudo conferir la capacidad de elevarse en el aire durante sus actividades depredadoras y, con el transcurso del tiempo, hizo posible que aquellas criaturas evolucionaran en el aire con vuelo a impulso propio.
Gerald Caple, un farmacéutico, encontró poco probable que la red para cazar insectos pudiera haber evolucionado hasta dar impulso elevatorio a las capacidades necesarias para el vuelo activo motor, mostraron cómo los corredores bípedos podían desarrollar la fuerza y la capacidad suficiente para volar.
Afirmaron que tales animales, corriendo y abalanzándose sobre sus presas, podrían haber utilizado sus extremidades anteriores para guardar el equilibrio, como hacen los seres humanos cuando dan un gran salto de longitud.
Esos movimientos de control se parecen rudimentariamente al golpe de ala en el vuelo. Si ese golpe de las extremidades superiores se llevaba a cabo cuando se daba un momento aerodinámicamente apto, como por ejemplo extremidades superiores cubiertas de plumas, el aire se deslizaría por encima y por debajo del ala a diferentes ritmos, con lo cual crearía un poder ascensional y prolongaría el salto.
Predijeron que el ritmo que llevaría al vuelo sería rápido. De acuerdo con sus hipótesis, el animal no necesitaba desarrollar músculos nuevos y diferentes para llevar a cabo el vuelo motor, en contraste con las necesidades de un planeador arbóreo.
Bock razonó de este modo: «Puesto que existen menos depredadores en los árboles que en el suelo, es posible que las protoaves se mantuvieran en los árboles mucho tiempo en condiciones que favorecían el desarrollo de una eficaz superficie aislante y, eventualmente, un cuerpo completamente cubierto de plumas. Más aún: si las protoaves utilizaban los árboles para anidar hubiera resultado muy ventajoso para los adultos de la especie el calentar los huevos en vez de dejarlos abandonados completamente para ser incubados por el calor solar.
Por eso las fuerzas de la selección natural en los árboles pudieron favorecer la evolución hacia la endotermia y el aislamiento mediante plumas».
Esta teoría, aunque muy poco probable, podría explicar otro aspecto intrigante de los dinosaurios y las aves:¿puede la sangre caliente de las aves ser presentada como evidencia en favor de que sus antepasados, los dinosaurios, también eran animales de sangre caliente?
Bock parece sugerir que los reptiles ya en camino de convertirse en aves desarrollaron su sangre caliente sólo en las etapas finales de su transición hacia las aves.
Robert Bakker ya había llegado a convencerse de que la endotermia no sólo era un rasgo común entre los dinosaurios, probablemente antes de su transición a las aves, sino que era una prueba convincente de la existencia de un estrecho parentesco dinosaurio-ave y, recíprocamente, que la sangre caliente de las aves era prueba evidente de que los dinosaurios fueron, también, animales endotérmicos.
Para Bakker el asignar a los dinosaurios una categoría superior resultaba algo lógico y justificado. Había entrado a saco en la paleontología con su declaración de la «superioridad» de los dinosaurios y ahora trataba de definir lo que en ellos había de especial en términos científicos.
Al igual que los mamíferos, arguyó, los dinosaurios habían dado un salto evolutivo trascendental con respecto a sus antepasados. Los mamíferos se habían desarrollado a partir de los reptiles, progresando desde la ectotermia a la endotermia y lo mismo habían hecho los dinosaurios. Además no tenía más sentido separar a las aves de los dinosaurios del que tendría separar a los murciélagos de los mamíferos.
Significa, como Ostrom escribiría en cierta ocasión, que los «dinosaurios no quedaron extinguidos sin dejar descendientes». Y que las plumas que crecieron en algunos dinosaurios no sólo llevaron al vuelo sino que fueron aislantes térmicos que a su vez fueron la razón primaria del éxito de los descendientes de los dinosaurios.
Predijeron que el ritmo que llevaría al vuelo sería rápido. De acuerdo con sus hipótesis, el animal no necesitaba desarrollar músculos nuevos y diferentes para llevar a cabo el vuelo motor, en contraste con las necesidades de un planeador arbóreo.
Bock razonó de este modo: «Puesto que existen menos depredadores en los árboles que en el suelo, es posible que las protoaves se mantuvieran en los árboles mucho tiempo en condiciones que favorecían el desarrollo de una eficaz superficie aislante y, eventualmente, un cuerpo completamente cubierto de plumas. Más aún: si las protoaves utilizaban los árboles para anidar hubiera resultado muy ventajoso para los adultos de la especie el calentar los huevos en vez de dejarlos abandonados completamente para ser incubados por el calor solar.
Por eso las fuerzas de la selección natural en los árboles pudieron favorecer la evolución hacia la endotermia y el aislamiento mediante plumas».
Esta teoría, aunque muy poco probable, podría explicar otro aspecto intrigante de los dinosaurios y las aves:¿puede la sangre caliente de las aves ser presentada como evidencia en favor de que sus antepasados, los dinosaurios, también eran animales de sangre caliente?
Bock parece sugerir que los reptiles ya en camino de convertirse en aves desarrollaron su sangre caliente sólo en las etapas finales de su transición hacia las aves.
Robert Bakker ya había llegado a convencerse de que la endotermia no sólo era un rasgo común entre los dinosaurios, probablemente antes de su transición a las aves, sino que era una prueba convincente de la existencia de un estrecho parentesco dinosaurio-ave y, recíprocamente, que la sangre caliente de las aves era prueba evidente de que los dinosaurios fueron, también, animales endotérmicos.
Para Bakker el asignar a los dinosaurios una categoría superior resultaba algo lógico y justificado. Había entrado a saco en la paleontología con su declaración de la «superioridad» de los dinosaurios y ahora trataba de definir lo que en ellos había de especial en términos científicos.
Al igual que los mamíferos, arguyó, los dinosaurios habían dado un salto evolutivo trascendental con respecto a sus antepasados. Los mamíferos se habían desarrollado a partir de los reptiles, progresando desde la ectotermia a la endotermia y lo mismo habían hecho los dinosaurios. Además no tenía más sentido separar a las aves de los dinosaurios del que tendría separar a los murciélagos de los mamíferos.
Significa, como Ostrom escribiría en cierta ocasión, que los «dinosaurios no quedaron extinguidos sin dejar descendientes». Y que las plumas que crecieron en algunos dinosaurios no sólo llevaron al vuelo sino que fueron aislantes térmicos que a su vez fueron la razón primaria del éxito de los descendientes de los dinosaurios.
John R. Horner y Robert Makela, ambos paleontólogos, se dedicaron a cavar en piedra arenosa de color marrón grisáceo. Al cabo de pocos días, trabajando bajo un sol ardiente y un fuerte viento, excavaron los restos de un nido de barro que contenía quince pequeñas crías de dinosaurios fosilizadas.
Cada una de las crías tenía aproximadamente un metro de longitud. Éstas fueron las primeras crías de dinosaurio encontradas en su nido y sus dientes mostraban que llevaban algún tiempo comiendo.
Eso señaló el comienzo de uno de los descubrimientos de dinosaurios más brillantes del siglo XX. Horner y Makela descubrieron más de trescientos huevos de dinosaurio en distintos nidos. Algunos huevos conservaban el esqueleto del embrión. Los dos buscadores sacaron más de sesenta esqueletos, totales o parciales, incluyendo los de las crías, y otros de dinosaurios muy jóvenes, así como los de sus padres.
Nada de lo que había sido desenterrado hasta entonces había aportado un material básico informativo para poder conocer la conducta social de los dinosaurios. Así se obtuvo la primera prueba evidente de que los dinosaurios se ocupaban del cuidado de sus crías, un rasgo característico que no se da en absoluto en los reptiles actuales.
Más tarde, en el verano de su primera temporada de trabajo, Laurie Trexler, que los estaba ayudando, encontró allí la cabeza de un dinosaurio adulto. «Si me tengo que conformar con una parte de un animal -explicó Horner-, ciertamente prefiero que sea el cráneo». Aquél le intrigó aún más, pues parecía distinto de todos los dinosaurios conocidos hasta entonces.
Durante meses Horner examinó el espécimen. Finalmente decidió que pertenecía a un miembro de la subfamilia de los arcosaurios conocida como hadrosauridae, el primero de los dinosaurios de pico de pato que fuera descubierto.
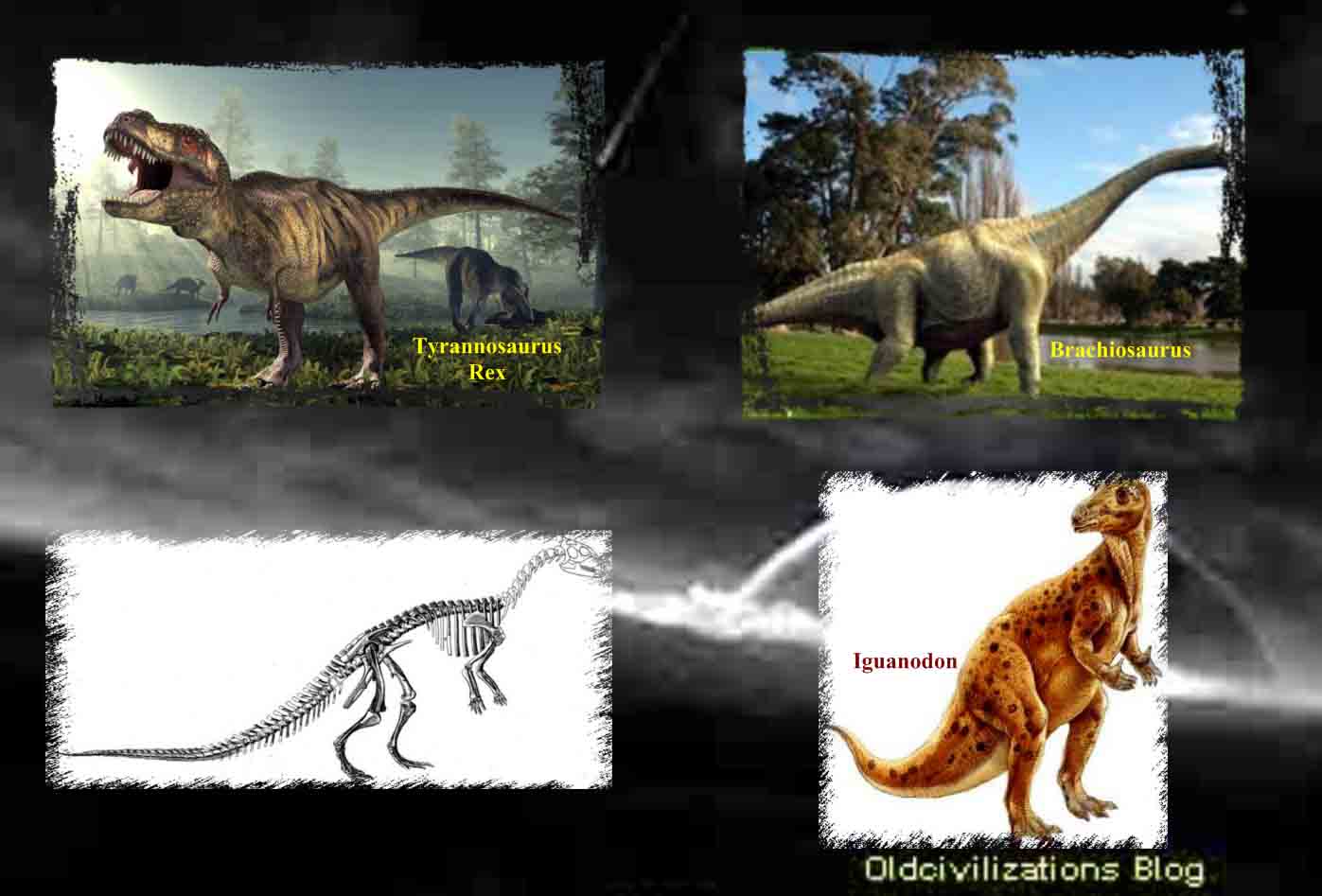
Eran los más abundantes y diversos de los dinosaurios del cretáceo. La mayor parte de los otros hadrosaurios tenían una abertura de las fosas nasales muy grande que incluía las órbitas de los ojos. Aquél, por el contrario, tenía unas fosas nasales pequeñas con un hueso muy grande entre éstas y las órbitas. A ese respecto, el animal se parecía al Iguanodon, pero éste tenía dientes pequeños, sencillos y escasos.
El cráneo que intrigaba a Horner tenía una verdadera batería de cientos de grandes dientes unidos entre sí y que eran mucho más adecuados para una mejor masticación y el roído de la vegetación dura. Esta característica y algunas otras sugerían que el animal debió de tener entre los ocho y los diez metros de longitud.
Finalmente llegó a la conclusión de que se trataba de un hadrosaurio de un nuevo género y especie, a los que bautizó con el nombre de Maiasaura peeblesorum. Maiasaura significa algo así como «la buena madre lagarto», nombre que parece muy apropiado a partir de la evidencia.
Cada una de las crías tenía aproximadamente un metro de longitud. Éstas fueron las primeras crías de dinosaurio encontradas en su nido y sus dientes mostraban que llevaban algún tiempo comiendo.
Eso señaló el comienzo de uno de los descubrimientos de dinosaurios más brillantes del siglo XX. Horner y Makela descubrieron más de trescientos huevos de dinosaurio en distintos nidos. Algunos huevos conservaban el esqueleto del embrión. Los dos buscadores sacaron más de sesenta esqueletos, totales o parciales, incluyendo los de las crías, y otros de dinosaurios muy jóvenes, así como los de sus padres.
Nada de lo que había sido desenterrado hasta entonces había aportado un material básico informativo para poder conocer la conducta social de los dinosaurios. Así se obtuvo la primera prueba evidente de que los dinosaurios se ocupaban del cuidado de sus crías, un rasgo característico que no se da en absoluto en los reptiles actuales.
Más tarde, en el verano de su primera temporada de trabajo, Laurie Trexler, que los estaba ayudando, encontró allí la cabeza de un dinosaurio adulto. «Si me tengo que conformar con una parte de un animal -explicó Horner-, ciertamente prefiero que sea el cráneo». Aquél le intrigó aún más, pues parecía distinto de todos los dinosaurios conocidos hasta entonces.
Durante meses Horner examinó el espécimen. Finalmente decidió que pertenecía a un miembro de la subfamilia de los arcosaurios conocida como hadrosauridae, el primero de los dinosaurios de pico de pato que fuera descubierto.
Eran los más abundantes y diversos de los dinosaurios del cretáceo. La mayor parte de los otros hadrosaurios tenían una abertura de las fosas nasales muy grande que incluía las órbitas de los ojos. Aquél, por el contrario, tenía unas fosas nasales pequeñas con un hueso muy grande entre éstas y las órbitas. A ese respecto, el animal se parecía al Iguanodon, pero éste tenía dientes pequeños, sencillos y escasos.
El cráneo que intrigaba a Horner tenía una verdadera batería de cientos de grandes dientes unidos entre sí y que eran mucho más adecuados para una mejor masticación y el roído de la vegetación dura. Esta característica y algunas otras sugerían que el animal debió de tener entre los ocho y los diez metros de longitud.
Finalmente llegó a la conclusión de que se trataba de un hadrosaurio de un nuevo género y especie, a los que bautizó con el nombre de Maiasaura peeblesorum. Maiasaura significa algo así como «la buena madre lagarto», nombre que parece muy apropiado a partir de la evidencia.
Horner y Makela continuaron buscando y excavando juntos en años sucesivos. De este modo pronto hicieron descubrimientos suficientes para darse cuenta de que no habían descubierto un nido aislado, o dos, sino toda una colonia.
Pronto excavaron dos nidos más con crías dehadrosaurios. En uno de los nidos las crías eran aún más jóvenes que los encontrados anteriormente. Medían menos de medio metro. Otras eran de más edad y habían crecido hasta los dos metros de longitud, lo que daba a Horner nuevos argumentos para creer que aquellas crías y pequeños eran cuidados y atendidos por sus padres durante varios meses después de la incubación y la ruptura del huevo.
El Maiasaura, «la buena madre lagarto», debió de haber alimentado a sus hijos, regresando frecuentemente al nido de fango arenoso con semillas, bayas y hojas verdes. El equipo de Horner también encontró los restos destrozados por el tiempo de nueve nidos vacíos, todos los cuales contenían gran abundancia de trozos de cáscaras de huevos fosilizadas.
Aquellos huevos de hadrosaurio, una vez fueron reconstruidos, medían unos veinte centímetros de longitud con la forma de elipsoides chatos, aproximadamente del tamaño y la forma de los huevos del Protoceratops que anteriormente habían sido hallados en el desierto de Gobi. Las cáscaras eran algo más gruesas que las de un huevo de gallina, más bien como las de un huevo de avestruz. Las superficies externas de las cáscaras eran rugosas.
Los nidos estaban separados entre sí unos siete metros como mínimo, lo que Horner observó con especial atención, porque siete metros suele ser, aproximadamente, la longitud del hadrosaurio adulto. Todos los nidos ocupaban la misma capa o estrato, lo que parecía indicar un mismo «horizonte temporal».
Eso hacía suponer que aquel lugar había sido sede de una colonia de crianza semejante a las de algunas aves modernas. En 1979, mientras cavaba en un montículo próximo, Horner descubrió otra colonia de nidos. Por esa razón el montículo fue bautizado con el nombre de Egg Mountain (montaña de los huevos).
Los diez nidos encontrados allí contenían los restos de nada menos que veinticuatro huevos cada uno. Aquellos huevos, con superficies exteriores lisas, eran algo más pequeños y más elipsoidales que los hallados anteriormente. Estaban colocados de modo circular con el extremo más puntiagudo enterrado en el fondo arenoso del nido.
Terminado el empollamiento la criatura salía por la parte superior del huevo, dejando tras sí la parte baja intacta entre los sedimentos. A partir de los huevos y de algunos huesos, Horner llegó a la conclusión que aquella colonia había sido ocupada por otra especie de dinosaurio, probablemente una especie del algo más pequeñoHypsilophodon.
Horner estuvo en condiciones de sacar dos deducciones a partir de la colonia de nidos de la Egg Mountain. Los nidos se encontraban a tres niveles diferentes sobre el suelo. Esto le sugería la idea de que los adultos habían ido anidando en el mismo lugar, año tras año, durante bastante tiempo. Es decir, era el lugar al que regresaban en cada temporada de crianza.
Pronto excavaron dos nidos más con crías dehadrosaurios. En uno de los nidos las crías eran aún más jóvenes que los encontrados anteriormente. Medían menos de medio metro. Otras eran de más edad y habían crecido hasta los dos metros de longitud, lo que daba a Horner nuevos argumentos para creer que aquellas crías y pequeños eran cuidados y atendidos por sus padres durante varios meses después de la incubación y la ruptura del huevo.
El Maiasaura, «la buena madre lagarto», debió de haber alimentado a sus hijos, regresando frecuentemente al nido de fango arenoso con semillas, bayas y hojas verdes. El equipo de Horner también encontró los restos destrozados por el tiempo de nueve nidos vacíos, todos los cuales contenían gran abundancia de trozos de cáscaras de huevos fosilizadas.
Aquellos huevos de hadrosaurio, una vez fueron reconstruidos, medían unos veinte centímetros de longitud con la forma de elipsoides chatos, aproximadamente del tamaño y la forma de los huevos del Protoceratops que anteriormente habían sido hallados en el desierto de Gobi. Las cáscaras eran algo más gruesas que las de un huevo de gallina, más bien como las de un huevo de avestruz. Las superficies externas de las cáscaras eran rugosas.
Los nidos estaban separados entre sí unos siete metros como mínimo, lo que Horner observó con especial atención, porque siete metros suele ser, aproximadamente, la longitud del hadrosaurio adulto. Todos los nidos ocupaban la misma capa o estrato, lo que parecía indicar un mismo «horizonte temporal».
Eso hacía suponer que aquel lugar había sido sede de una colonia de crianza semejante a las de algunas aves modernas. En 1979, mientras cavaba en un montículo próximo, Horner descubrió otra colonia de nidos. Por esa razón el montículo fue bautizado con el nombre de Egg Mountain (montaña de los huevos).
Los diez nidos encontrados allí contenían los restos de nada menos que veinticuatro huevos cada uno. Aquellos huevos, con superficies exteriores lisas, eran algo más pequeños y más elipsoidales que los hallados anteriormente. Estaban colocados de modo circular con el extremo más puntiagudo enterrado en el fondo arenoso del nido.
Terminado el empollamiento la criatura salía por la parte superior del huevo, dejando tras sí la parte baja intacta entre los sedimentos. A partir de los huevos y de algunos huesos, Horner llegó a la conclusión que aquella colonia había sido ocupada por otra especie de dinosaurio, probablemente una especie del algo más pequeñoHypsilophodon.
Horner estuvo en condiciones de sacar dos deducciones a partir de la colonia de nidos de la Egg Mountain. Los nidos se encontraban a tres niveles diferentes sobre el suelo. Esto le sugería la idea de que los adultos habían ido anidando en el mismo lugar, año tras año, durante bastante tiempo. Es decir, era el lugar al que regresaban en cada temporada de crianza.
Al advertir la ausencia de restos de jóvenes en aquellos nidos, así como el hecho de que las partes inferiores de los huevos estaban bien conservadas y no rotas como en los nidos del anterior descubrimiento, sin duda a causa del pataleo y movimiento de los pequeños, Horner dedujo que los jóvenes de esta especie no se quedaban en el nido tanto tiempo como los de la otra y lo abandonaban pronto tras haber salido del huevo.
No obstante había signos que indicaban la atención de los padres. Restos de entre quince y veinte esqueletos pequeños, de longitudes que iban desde medio metro a un metro y medio fueron encontrados unos cerca de otros.
Si las crías abandonaban el nido inmediatamente después de salir del cascarón, es posible que por casualidad algunos hubieran muerto allí, pero un grupo de quince o veinte parecía poco probable. Esa circunstancia recordó a Horner los lugares próximos al nido en los que se desarrollan algunas aves de nuestro tiempo.
Como informó en Nature, aquellos jóvenes dinosaurios «o bien seguían en la colonia o regresaban al lugar frecuentemente». Ambas cosas hubieran sido indicios de la existencia del cuidado paternal. Los análisis del desarrollo de los huesos de las crías de dinosaurios llevaron a Horner a nuevas evidencias. Descubrió la existencia de un crecimiento rápido.
El metabolismo de aquellas criaturas parecía haber tenido el ritmo rápido de los animales de sangre caliente más que el lento y perezoso de los reptiles de sangre fría. Su razonamiento era que los nidos de los hadrosauríos, en particular, contenían crías en los cascarones que ni siquiera llegaban a medir medio metro, así como restos de animales muy jóvenes de un metro y fragmentos de otros que podrían haber llegado a los dos metros. O bien crecían muy rápidamente o seguían mucho tiempo en el nido.
El cocodrilo joven, que es un animal de sangre fría, así como el arcosaurío, crecen un tercio de metro por año. Incluso las especies de más rápido crecimiento entre los animales de sangre fría tardarían un año en conseguir el tamaño de los jóvenes hadrosauríos descubiertos por Horner. Parece improbable que los jóvenes dinosaurios se quedaran en el nido tres años, o incluso uno, por muy cariñosos y dispuestos a alimentarlos que estuvieran sus padres.
Más bien habría que pensar que los dinosaurios crecían a un ritmo comparable al de los avestruces, por ejemplo, que es la criatura de sangre caliente de más lento crecimiento. Un avestruz alcanza un tamaño de un metro en un período de entre seis y ocho meses.
Por alguna razón las crías de la buena madre lagarto de Montana parecían capaces de crecer a ese ritmo o incluso a otro más rápido y esto podría indicar que se trataba de animales de sangre caliente.
No obstante había signos que indicaban la atención de los padres. Restos de entre quince y veinte esqueletos pequeños, de longitudes que iban desde medio metro a un metro y medio fueron encontrados unos cerca de otros.
Si las crías abandonaban el nido inmediatamente después de salir del cascarón, es posible que por casualidad algunos hubieran muerto allí, pero un grupo de quince o veinte parecía poco probable. Esa circunstancia recordó a Horner los lugares próximos al nido en los que se desarrollan algunas aves de nuestro tiempo.
Como informó en Nature, aquellos jóvenes dinosaurios «o bien seguían en la colonia o regresaban al lugar frecuentemente». Ambas cosas hubieran sido indicios de la existencia del cuidado paternal. Los análisis del desarrollo de los huesos de las crías de dinosaurios llevaron a Horner a nuevas evidencias. Descubrió la existencia de un crecimiento rápido.
El metabolismo de aquellas criaturas parecía haber tenido el ritmo rápido de los animales de sangre caliente más que el lento y perezoso de los reptiles de sangre fría. Su razonamiento era que los nidos de los hadrosauríos, en particular, contenían crías en los cascarones que ni siquiera llegaban a medir medio metro, así como restos de animales muy jóvenes de un metro y fragmentos de otros que podrían haber llegado a los dos metros. O bien crecían muy rápidamente o seguían mucho tiempo en el nido.
El cocodrilo joven, que es un animal de sangre fría, así como el arcosaurío, crecen un tercio de metro por año. Incluso las especies de más rápido crecimiento entre los animales de sangre fría tardarían un año en conseguir el tamaño de los jóvenes hadrosauríos descubiertos por Horner. Parece improbable que los jóvenes dinosaurios se quedaran en el nido tres años, o incluso uno, por muy cariñosos y dispuestos a alimentarlos que estuvieran sus padres.
Más bien habría que pensar que los dinosaurios crecían a un ritmo comparable al de los avestruces, por ejemplo, que es la criatura de sangre caliente de más lento crecimiento. Un avestruz alcanza un tamaño de un metro en un período de entre seis y ocho meses.
Por alguna razón las crías de la buena madre lagarto de Montana parecían capaces de crecer a ese ritmo o incluso a otro más rápido y esto podría indicar que se trataba de animales de sangre caliente.

Casi todos sus hallazgos de huesos habían sido hechos en tales sedimentos.
Sin embargo, Horner encontró sus nidos en terrenos que fueron más elevados y más áridos. De acuerdo con la hipótesis de Horner los dinosaurios emigraron de las tierras bajas, situadas a unos cien kilómetros, para hacer sus nidos, poner sus huevos y cuidar a sus crías en lugar alejado y seguro, que protegiera a sus hijos de los dinosaurios carnívoros.
Los primeros aspectos de la forma de vida de los dinosaurios que pudieron ser deducidos con cierto grado de certeza, como hemos podido ver, se relacionan con la locomoción y la manutención. Si las extremidades delanteras del animal habían sido considerablemente más pequeñas que sus miembros traseros, podía presumirse que esto los hacía bípedos.
Esto podía ser cierto si los miembros anteriores terminaban en manos con garras afiladas para agarrar y atacar. Pero algunos dinosaurios tenían unos crecimientos óseos, parecidos a cascos, en los dedos de los miembros más cortos y, por lo tanto, eran cuadrúpedos, o al menos lo fueron durante algún tiempo. Los grandes saurópodos, con cuatro miembros fuertes, como columnas y patas de elefante, eran indudablemente cuadrúpedos.
Los rastros de sus huellas fosilizadas lo confirman. La forma de los huesos de las extremidades y la configuración de las caderas y los hombros, por otra parte, prueban que los dinosaurios se alzaban más rectos y erguidos que otros reptiles que mantienen su postura clásica de animales reptantes. Los dientes también nos ofrecen claras indicaciones que nos ayudan a establecer su dieta. Si los animales tenían pequeños dientes más bien planos esto indicaba que se alimentaban de vegetales blandos.
Si los dientes eran mayores y estaban situados en las mandíbulas para trabajar en la masticación como si fueran tijeras, los animales podrían haber comido vegetación más dura y fibrosa que requería ser cortada y triturada. Algunos dinosaurios no tenían en absoluto dientes, lo que indica que subsistían alimentándose de insectos, huevos y frutas muy blandas.
Aquellos otros con los dientes más amenazadores, largos, agudos y en forma de sierra, eran carnívoros que cazaban a los herbívoros mientras éstos pastaban, lo mismo que los leones cazan en la actualidad. Los huesos delatan también los gustos alimenticios de los dinosaurios. Los huesos más densos y laminados pertenecen a los carnívoros.
En algunos pocos casos los científicos han encontrado pruebas directas de la dieta de un dinosaurio. Fue hallado un anatosaurio momificado que murió con el estómago lleno y su contenido se componía de agujas de coniferas, ramas, frutos y semillas.
Es posible que algunos dinosaurios ingirieran rocas, como algunas aves tragan finos guijarros para ayudarse en su digestión. Los cazadores de esqueletos han encontrado frecuentemente piedras extremadamente suaves, mezcladas con los esqueletos de los saurópodos.
Se las llama gastrolitos (piedras gástricas). Lo más probable es que esas piedras fueran tan finamente pulidas por las aguas de algún riachuelo existente en otros tiempos y no por los jugos gástricos de los dinosaurios.
A lo largo de los años los científicos han intentado obtener cierta idea sobre la fisiología y la conducta del dinosaurio basándose en el tamaño y la forma de sus cerebros. Sus cerebros eran extremadamente pequeños en relación con sus cuerpos y consecuentemente, su inteligencia no debió de ser muy alta.
Pero algunos de sus sentidos podrían compensar algo de esta deficiencia mental. Un cuidadoso examen de la estructura cerebral ha revelado la existencia de bulbos olfativos y lóbulos ópticos bien desarrollados, lo que indica una buena capacidad olfativa y de visión.
La estructura ósea interna de las orejas sugiere un excelente oído. Posiblemente estaban capacitados para oír notas extremadamente altas, quizá los agudos grítitos de sus crías, una habilidad muy importante para mantener la cohesión familiar.
Todos los dinosaurios tienen órbitas demasiado grandes para sus ojos. ¿De qué color? Nadie lo sabe, pero los actuales reptiles tienen ojos de un color que va del rojo al amarillo. No hay ninguna prueba de que los dinosaurios pudieran expresar oralmente los pensamientos que pudieran cruzar por sus pequeños cerebros.
Pero sí hay presunciones en ese sentido. Algunos huesos de sus cráneos sugieren que tenían una voz potente. W. E. Swinton, un paleontólogo británico, ha escrito: «En este aspecto podrían haber igualado a los cocodrilos, todos los cuales tienen un corto y agudo croar, o una especie de ladrido, mediante el cual se identifican entre sí en la oscuridad o que utilizan para indicar cuándo están irritados». Los dinosaurios, como todos los animales, debían de tener sus medios de ataque, de defensa o de huida ante los ataques.
Los herbívoros que pastaban en las llanuras tenían que confiar en su vista y en su oído para descubrir a los depredadores y poder escapar, al tiempo que croaban, ladraban o dejaban escapar su sonido de tuba, para alertar a los otros del inminente peligro.
Otros contaban para defenderse con sus garras, sus aguzados espolones, sus colas que utilizaban como trancas y sus cuernos aguzados, útiles tanto para el ataque como para la defensa. Los Triceratopsposiblemente luchaban entre sí como los cameros, empujándose y golpeándose con sus cráneos y sus fuertes cuernos. Sus esqueletos a veces mostraban señales de profundas heridas en la coraza ósea que se proyectaba hacia atrás como un escudo sobre el cuello y los hombros.
Se las llama gastrolitos (piedras gástricas). Lo más probable es que esas piedras fueran tan finamente pulidas por las aguas de algún riachuelo existente en otros tiempos y no por los jugos gástricos de los dinosaurios.
A lo largo de los años los científicos han intentado obtener cierta idea sobre la fisiología y la conducta del dinosaurio basándose en el tamaño y la forma de sus cerebros. Sus cerebros eran extremadamente pequeños en relación con sus cuerpos y consecuentemente, su inteligencia no debió de ser muy alta.
Pero algunos de sus sentidos podrían compensar algo de esta deficiencia mental. Un cuidadoso examen de la estructura cerebral ha revelado la existencia de bulbos olfativos y lóbulos ópticos bien desarrollados, lo que indica una buena capacidad olfativa y de visión.
La estructura ósea interna de las orejas sugiere un excelente oído. Posiblemente estaban capacitados para oír notas extremadamente altas, quizá los agudos grítitos de sus crías, una habilidad muy importante para mantener la cohesión familiar.
Todos los dinosaurios tienen órbitas demasiado grandes para sus ojos. ¿De qué color? Nadie lo sabe, pero los actuales reptiles tienen ojos de un color que va del rojo al amarillo. No hay ninguna prueba de que los dinosaurios pudieran expresar oralmente los pensamientos que pudieran cruzar por sus pequeños cerebros.
Pero sí hay presunciones en ese sentido. Algunos huesos de sus cráneos sugieren que tenían una voz potente. W. E. Swinton, un paleontólogo británico, ha escrito: «En este aspecto podrían haber igualado a los cocodrilos, todos los cuales tienen un corto y agudo croar, o una especie de ladrido, mediante el cual se identifican entre sí en la oscuridad o que utilizan para indicar cuándo están irritados». Los dinosaurios, como todos los animales, debían de tener sus medios de ataque, de defensa o de huida ante los ataques.
Los herbívoros que pastaban en las llanuras tenían que confiar en su vista y en su oído para descubrir a los depredadores y poder escapar, al tiempo que croaban, ladraban o dejaban escapar su sonido de tuba, para alertar a los otros del inminente peligro.
Otros contaban para defenderse con sus garras, sus aguzados espolones, sus colas que utilizaban como trancas y sus cuernos aguzados, útiles tanto para el ataque como para la defensa. Los Triceratopsposiblemente luchaban entre sí como los cameros, empujándose y golpeándose con sus cráneos y sus fuertes cuernos. Sus esqueletos a veces mostraban señales de profundas heridas en la coraza ósea que se proyectaba hacia atrás como un escudo sobre el cuello y los hombros.
En 1971 la expedición polaco-mongola encontró en Gobi los restos de dos dinosaurios que, posiblemente, se habían matado entre sí. El depredador, un pequeño y rápido Velociraptor; estaba abrazado sobre su presa, un Protoceratops acorazado.
El Velociraptor tenía un espolón en forma de hoz y muy afilado en cada una de sus patas traseras, que se parecía mucho al delDeinonychus de Ostrom. Uno de los espolones del depredador estaba hundido en la región que debió de ser el vientre de la presa. No estaba claro cómo el Protoceratops había logrado su póstuma venganza.
Sin embargo este descubrimiento fue considerado como una prueba más de la nueva teoría que consideraba a los dinosaurios como más ágiles y activos que los reptiles ordinarios. Más aún se ha podido deducir de las huellas dejadas por los dinosaurios. Las huellas de las patas, como en cierta ocasión diría Richard Swann, «son fósiles de seres vivos, mientras que todos los demás fósiles son reliquias de los muertos».
De las series de huellas de patas los científicos han deducido la velocidad de los dinosaurios corredores, han comprobado cómo los depredadores perseguían a sus presas, los saurópodos, deduciendo datos que los han llevado a su primera comprensión de la conducta en grupo de los dinosaurios.
Éstos, al parecer, viajaban frecuentemente en rebaños, por lo cual podrían ser animales gregarios. Uno de los descubrimientos más importantes debidos a las huellas rescata en cierto sentido al Brontosaurusdel agua, y lo sitúa en tierra firme.
Algunos libros ilustrados aún representan a los Brontosauruscon sus hocicos en aguas pantanosas, en actitud letárgica, casi inmóvil, masticando plantas acuáticas blandas. Los paleontólogos parecían creer que éste era el papel reservado en la vida a los Brontosaurus.
Sus patas no eran lo suficientemente robustas para servir de apoyo a sus pesados cuerpos, alegaban los científicos, y por lo tanto necesitaban la flotabilidad de sus cuerpos dentro del agua. Además estaba la abertura nasal en la parte superior de la cabeza, lo que al parecer permitía respirar al animal aun con su cuerpo completamente sumergido.
Todo esto dio lugar a la imagen lenta y pesada del Brontosaurus presentándolo como una especie de hipopótamo de los dinosaurios, aunque aún más lento y perezoso. La imagen comenzó a cambiar -en la mente de los científicos después de que Roland T. Bird, del Museo Americano de Historia Natural, examinó algunas huellas de dinosaurios en Texas, en la década de los años 1940-1950. Bird dio con la secuencia de huellas del mismo modo que Horner encontró las crías de dinosaurios.
El Velociraptor tenía un espolón en forma de hoz y muy afilado en cada una de sus patas traseras, que se parecía mucho al delDeinonychus de Ostrom. Uno de los espolones del depredador estaba hundido en la región que debió de ser el vientre de la presa. No estaba claro cómo el Protoceratops había logrado su póstuma venganza.
Sin embargo este descubrimiento fue considerado como una prueba más de la nueva teoría que consideraba a los dinosaurios como más ágiles y activos que los reptiles ordinarios. Más aún se ha podido deducir de las huellas dejadas por los dinosaurios. Las huellas de las patas, como en cierta ocasión diría Richard Swann, «son fósiles de seres vivos, mientras que todos los demás fósiles son reliquias de los muertos».
De las series de huellas de patas los científicos han deducido la velocidad de los dinosaurios corredores, han comprobado cómo los depredadores perseguían a sus presas, los saurópodos, deduciendo datos que los han llevado a su primera comprensión de la conducta en grupo de los dinosaurios.
Éstos, al parecer, viajaban frecuentemente en rebaños, por lo cual podrían ser animales gregarios. Uno de los descubrimientos más importantes debidos a las huellas rescata en cierto sentido al Brontosaurusdel agua, y lo sitúa en tierra firme.
Algunos libros ilustrados aún representan a los Brontosauruscon sus hocicos en aguas pantanosas, en actitud letárgica, casi inmóvil, masticando plantas acuáticas blandas. Los paleontólogos parecían creer que éste era el papel reservado en la vida a los Brontosaurus.
Sus patas no eran lo suficientemente robustas para servir de apoyo a sus pesados cuerpos, alegaban los científicos, y por lo tanto necesitaban la flotabilidad de sus cuerpos dentro del agua. Además estaba la abertura nasal en la parte superior de la cabeza, lo que al parecer permitía respirar al animal aun con su cuerpo completamente sumergido.
Todo esto dio lugar a la imagen lenta y pesada del Brontosaurus presentándolo como una especie de hipopótamo de los dinosaurios, aunque aún más lento y perezoso. La imagen comenzó a cambiar -en la mente de los científicos después de que Roland T. Bird, del Museo Americano de Historia Natural, examinó algunas huellas de dinosaurios en Texas, en la década de los años 1940-1950. Bird dio con la secuencia de huellas del mismo modo que Horner encontró las crías de dinosaurios.
Cada una de las huellas tenía un metro de longitud y estaban impresas con fuerza, profundamente, en lo que debió de ser una superficie fangosa parcial o totalmente expuesta al aire.
Las huellas, separadas entre sí por dos metros, eran definitivamente las de un animal terrestre. El Brontosaurus podía andar sobre sus cuatro patas. En otro lugar de Texas, Bird vio una continua sucesión de huellas de las patas del Brontosaurus en la roca.
El animal había arrastrado su pesada cola sobre la superficie fangosa, otra señal de que andaba y no nadaba o flotaba a medias en el agua. De estas pruebas los paleontólogos dedujeron algo que ya hacía mucho tiempo que debieron saber: que el Brontosaurus no era un animal prisionero del agua.
Sus fósiles no fueron hallados, según pudo comprobarse, en sedimentos de origen pantanoso, sino en los mismos sedimentos procedentes de aluviones que contienen los restos de los demás dinosaurios que están clasificados como animales terrestres.
El estudio de las huellas fósiles es una rama de la paleontología que se conoce con el nombre de ichnografia, derivado de la palabra griega que significa huella o rastro. Las primeras sendas marcadas por las huellas de los dinosaurios que llamaron la atención de modo más general fueron las impresiones de patas parecidas a las de las aves descubiertas en el valle del río Connecticut en Massachusetts, en 1802, cuando los dinosaurios aún eran desconocidos. Desde entonces han sido descubiertos cientos de otras series de huellas en Nueva Inglaterra, Texas, Brasil, Australia y, más recientemente, en el cañón del río Peace en la Columbia Británica.
Naturalmente las huellas de los dinosaurios son más abundantes que los huesos de dinosaurio. Mediante un análisis realizado por los científicos en las huellas, utilizando una fórmula establecida por R. McNeill Alexander de la Universidad de Leed, determinaron que algunos dinosaurios eran capaces de moverse con bastante rapidez. Algunos carnívoros de tamaño medio marchaban a una media de 16,5 kilómetros por hora, lo cual se aproxima a la máxima velocidad de carrera conseguida por el ser humano.
Algunos de los carnívoros más lentos lo hacían entre los 6 y los 8,5 kilómetros por hora. Los herbívoros incluso podían ser más lentos con velocidades máximas de 6 kilómetros por hora. Con estos cálculos un ex discípulo de Ostrom, James O. Farlow, especialista en dinosaurios independiente, de Michigan, informó en 1981 que algunos carnívoros habían dejado huellas en Texas que demostraban que alcanzaban una velocidad de 42 kilómetros por hora, es decir, más rápidos que los seres humanos, pero que no podían competir con los caballos de buena raza ni con los galgos. Otros cálculos indicaban que en caso necesario el Tyrannosaurus podía superar los 45 o 50 kilómetros por hora.
Según se deduce de las huellas, los Tyrannosaurus solían marchar solos o en parejas. Pero el instinto de rebaño o manada debió de ser más fuerte en otras especies. En el Parque Estatal de Dinosaurios de Connecticut, muchas de las huellas transcurren paralelas como si los dinosaurios marcharan en rebaño.
En las sendas dejadas por los saurópodos en Texas y descubiertas por Roland Bird también existen pruebas de que marchaban en rebaño. Bakker en un reciente reexamen de esas huellas cree haber reconocido señales de un rebaño «estructurado», con los más jóvenes y las crías en el centro rodeados y protegidos por los adultos.
En Montana, Jack Horner encontró pruebas de que sus hadrosauríos no sólo constituían colonias ponedoras sino también que continuaban constituidos en grupo durante la mayor parte, o la totalidad, de su vida.
En varios lugares encontró restos de jóvenes de unos tres metros de longitud con restos de adultos de seis o siete metros. En un lugar había veinticuatro jóvenes junto a diez adultos.
Es posible que no parezca demasiado importante el saber que algunos dinosaurios viajaban en manadas o en rebaños y que otros cuidaban a sus crías, pero lo cierto es que esa conducta los diferencia claramente de otros reptiles. La mayor parte de los reptiles, como los peces y los anfibios, abandonan sus huevos tan pronto como han sido puestos.
Algunos peces y reptiles, entre ellos el cocodrilo, guardan sus huevos hasta que termina la incubación o un poco después y una vez nacidas las crías tienen que alimentarse y cuidarse por sí solas. Ninguno de los reptiles modernos vive en rebaños o en manadas de caza ni en ningún otro tipo de comunidad social. Los dinosaurios fueron diferentes.
Algunos de ellos tenían vida familiar y un sentido de comunidad. Horner creía que la conducta social que podía inferirse de las huellas y los nidos podía ser de gran importancia en el éxito tremendo de los dinosaurios que hizo posible que su existencia se prolongara durante ciento sesenta millones de años.
Los sociobiólogos creen que ciertos comportamientos fundamentales pasan de una generación a otra por medio de los genes. Uno de esos rasgos, el altruismo en el reparto de los alimentos, se observa de manera primordial en las aves y en los mamíferos.
Si los padres delMaiasaura llevaban la comida a sus crías en el nido, como Horner creía, era posible que su especie y algunas otras especies de dinosaurios se transmitieran el altruismo de modo genético. Sin ese altruismo y cierto instinto social, en el cual se incluía el reparto de la comida, era más que posible que un buen número de dinosaurios no hubiera podido llevar una vida próspera en un mundo lleno de depredadores.
En las sendas dejadas por los saurópodos en Texas y descubiertas por Roland Bird también existen pruebas de que marchaban en rebaño. Bakker en un reciente reexamen de esas huellas cree haber reconocido señales de un rebaño «estructurado», con los más jóvenes y las crías en el centro rodeados y protegidos por los adultos.
En Montana, Jack Horner encontró pruebas de que sus hadrosauríos no sólo constituían colonias ponedoras sino también que continuaban constituidos en grupo durante la mayor parte, o la totalidad, de su vida.
En varios lugares encontró restos de jóvenes de unos tres metros de longitud con restos de adultos de seis o siete metros. En un lugar había veinticuatro jóvenes junto a diez adultos.
Es posible que no parezca demasiado importante el saber que algunos dinosaurios viajaban en manadas o en rebaños y que otros cuidaban a sus crías, pero lo cierto es que esa conducta los diferencia claramente de otros reptiles. La mayor parte de los reptiles, como los peces y los anfibios, abandonan sus huevos tan pronto como han sido puestos.
Algunos peces y reptiles, entre ellos el cocodrilo, guardan sus huevos hasta que termina la incubación o un poco después y una vez nacidas las crías tienen que alimentarse y cuidarse por sí solas. Ninguno de los reptiles modernos vive en rebaños o en manadas de caza ni en ningún otro tipo de comunidad social. Los dinosaurios fueron diferentes.
Algunos de ellos tenían vida familiar y un sentido de comunidad. Horner creía que la conducta social que podía inferirse de las huellas y los nidos podía ser de gran importancia en el éxito tremendo de los dinosaurios que hizo posible que su existencia se prolongara durante ciento sesenta millones de años.
Los sociobiólogos creen que ciertos comportamientos fundamentales pasan de una generación a otra por medio de los genes. Uno de esos rasgos, el altruismo en el reparto de los alimentos, se observa de manera primordial en las aves y en los mamíferos.
Si los padres delMaiasaura llevaban la comida a sus crías en el nido, como Horner creía, era posible que su especie y algunas otras especies de dinosaurios se transmitieran el altruismo de modo genético. Sin ese altruismo y cierto instinto social, en el cual se incluía el reparto de la comida, era más que posible que un buen número de dinosaurios no hubiera podido llevar una vida próspera en un mundo lleno de depredadores.
Los hadrosaurios, por ejemplo, eran herbívoros bípedos que no disponían de ningún medio de defensa obvio, ya que carecían de cuernos, de garras o de protección corporal. Tampoco eran capaces de correr a gran velocidad, puesto que eran caminantes lentos.
Debieron de ser muy vulnerables ante el ataque de los depredadores, sobre todo cuando estaban solos. Consecuentemente, y como las nuevas pruebas parecían indicar, se vieron obligados a formar rebaños y a cuidar de sus crías durante varios meses después de que salieran de su cascarón, guardándolos en colonias relativamente bien protegidas.
Y debieron hacerlo bien puesto que pese a su vulnerabilidad los hadrosaurios se cuentan entre los dinosaurios más abundantes que existieron. Estaba claro que sobrevivía el número suficiente de jóvenes hadrosaurios para reemplazar a los adultos y mantener la población.
Según dijo Horner en 1983 esto fue así porque probablemente supieron emplear una «estrategia de supervivencia comparable a la más común entre las aves, la construcción de colonias de anidamiento». Ponían menos huevos que otros reptiles, pero protegían sus nidos durante la incubación y se dedicaban a alimentar a sus crías.
La anidación en colonia y la protección de los nidos es practicada por los cocodrilos, pero éstos no alimentan a sus crías ni comparten los alimentos. Para los hadrosaurios, argüía Horner, ambas cosas se hacían necesarias pues de otro modo hubiera sido más que cuestionable la supervivencia de una o dos de esas pequeñas y pacíficas criaturas de cada camada de veinte o veinticinco.
El hecho de que en sus grupos o rebaños hubiera al menos dos jóvenes por cada adulto sugiere una supervivencia del ocho por ciento de cada camada. El hecho de que en los nidos se encontrara un número tan grande de crías sugiere que eran los padres quienes se cuidaban de alimentarlos.
El salir de sus nidos para alimentarse por su cuenta habría significado para las crías el riesgo de ser devoradas por los depredadores o ser pisoteadas por los adultos. Como muchas aves, los pequeños hadrosaurios permanecían en sus nidos por instinto.
No sabemos durante cuánto tiempo, pero debió de ser durante varios meses, hasta alcanzar una longitud de metro y medio como mínimo. Aquellas quince crías encontradas por Horner en su primer nido debieron de quedarse en él mientras sus padres iban a buscarles comida. Sus padres no regresaron a causa de cualquier desastre, pero las crías se quedaron en el nido hasta que les sobrevino la muerte por inanición.
Es posible que esa conducta de cuidado paternal y compartir los alimentos estuviera restringida a sólo unas pocas especies, concluye Horner, pero en todo caso refleja una amplia «divergencia sociobiológica» entre las dos grandes ramas de los dinosaurios, los omitichianos y los saurischianos.
Debieron de ser muy vulnerables ante el ataque de los depredadores, sobre todo cuando estaban solos. Consecuentemente, y como las nuevas pruebas parecían indicar, se vieron obligados a formar rebaños y a cuidar de sus crías durante varios meses después de que salieran de su cascarón, guardándolos en colonias relativamente bien protegidas.
Y debieron hacerlo bien puesto que pese a su vulnerabilidad los hadrosaurios se cuentan entre los dinosaurios más abundantes que existieron. Estaba claro que sobrevivía el número suficiente de jóvenes hadrosaurios para reemplazar a los adultos y mantener la población.
Según dijo Horner en 1983 esto fue así porque probablemente supieron emplear una «estrategia de supervivencia comparable a la más común entre las aves, la construcción de colonias de anidamiento». Ponían menos huevos que otros reptiles, pero protegían sus nidos durante la incubación y se dedicaban a alimentar a sus crías.
La anidación en colonia y la protección de los nidos es practicada por los cocodrilos, pero éstos no alimentan a sus crías ni comparten los alimentos. Para los hadrosaurios, argüía Horner, ambas cosas se hacían necesarias pues de otro modo hubiera sido más que cuestionable la supervivencia de una o dos de esas pequeñas y pacíficas criaturas de cada camada de veinte o veinticinco.
El hecho de que en sus grupos o rebaños hubiera al menos dos jóvenes por cada adulto sugiere una supervivencia del ocho por ciento de cada camada. El hecho de que en los nidos se encontrara un número tan grande de crías sugiere que eran los padres quienes se cuidaban de alimentarlos.
El salir de sus nidos para alimentarse por su cuenta habría significado para las crías el riesgo de ser devoradas por los depredadores o ser pisoteadas por los adultos. Como muchas aves, los pequeños hadrosaurios permanecían en sus nidos por instinto.
No sabemos durante cuánto tiempo, pero debió de ser durante varios meses, hasta alcanzar una longitud de metro y medio como mínimo. Aquellas quince crías encontradas por Horner en su primer nido debieron de quedarse en él mientras sus padres iban a buscarles comida. Sus padres no regresaron a causa de cualquier desastre, pero las crías se quedaron en el nido hasta que les sobrevino la muerte por inanición.
Es posible que esa conducta de cuidado paternal y compartir los alimentos estuviera restringida a sólo unas pocas especies, concluye Horner, pero en todo caso refleja una amplia «divergencia sociobiológica» entre las dos grandes ramas de los dinosaurios, los omitichianos y los saurischianos.
 Otra especie de la que se sabe que ponía sus huevos en nidos circulares con una notable atención, como vio Roy Chapman Andrews, era la de los Protoceratops, también omitichianos.
Otra especie de la que se sabe que ponía sus huevos en nidos circulares con una notable atención, como vio Roy Chapman Andrews, era la de los Protoceratops, también omitichianos.Los huevos redondos de un dinosaurio parecido al Brontosaurus, quizá un Hypselosaurus, eran puestos en largas filas lineales que no es la mejor forma si se piensa en alimentar a las crías en el nido.
Podría ser, según dijo Horner, que los omitichianos pusieran sus huevos en círculo y lossaurischianos en filas. La conducta en cierto modo más sofisticada y social de algunos dinosaurios puede haber sido una adaptación evolucionaría relacionada con el herbivorismo.
Aunque es posible que unos pocos carnívoros cazaran en manada, los dinosaurios que según se sabe se desplazaban formando rebaños eran saurópodos herbívoros. «Incluso si se trataba sólo de reuniones y no de rebaños en el sentido estricto del término, se trata de un comportamiento que no vemos en otros reptiles -afirmó Horner-.
Eso es algo que sólo conocemos en los mamíferos y en las aves». Y lo que es más, subraya Horner, los omitichianosfueron «el primer gran grupo de herbívoros que habitó la Tierra y para poder sobrevivir, competir y muy especialmente lograr un éxito tan considerable, tuvieron necesidad de establecer un orden social fuerte».
Pero algo ocurrió hace sesenta y cinco millones de años, a finales del cretáceo, algo tan devastador que alteró el curso de la vida en la Tierra. De modo aparentemente repentino, en relación con el transcurrir del tiempo geológico, casi la mitad de los géneros vivientes desaparecieron. Ninguna de las especies que vivían exclusivamente en tierra y que pesaban más de veinticinco kilos parecen haber sobrevivido.
Y los más notables de los desaparecidos fueron los dinosaurios. Aunque los fósiles de una variedad de dinosaurios han sido encontrados en las capas superiores de las rocas del cretáceo, ni uno solo ha sido desenterrado de las capas del terciario que están inmediatamente encima. Los dinosaurios habían desaparecido para no volver a ser vistos jamás.
Las extinciones a finales del cretáceo hicieron caer el telón de la Era de los Reptiles. Pero muchos dinosaurios habían muerto con anterioridad. El Stegosaurus, de piel blindada, se extinguió antes de que comenzara el cretáceo. Los gigantescos Diplodocus, Brachiosaurus y Brontosaurus se extinguieron en el transcurso del cretáceo antes de que se produjera la gran agonía.
Todos o casi todos lossaurópodos de ese tipo habían desaparecido. La mayor parte de los iguanodontes se había extinguido para entonces, pero los dinosaurios aún existían hacia finales del cretáceo. Entre otros arcosaurios, los cocodrilos marinos y muchos ictiosaurios posiblemente se habían extinguido ya, pero los plesiosáuridos, los pterosaurios y los mosasáuridos parecen haber tenido su período de florecimiento a finales del cretáceo y después se extinguieron. Sólo algunos crocodílidos, entre todos los arcosaurios, se las arreglaron para cruzar esa frontera tan plena en acontecimientos que separa el cretáceo del terciario y han sobrevivido hasta nuestros días.
Ese algo que ocurrió hace sesenta y cinco millones de años es uno de los misterios más intrigantes de la ciencia. No obstante, los dinosaurios tuvieron una larga y próspera estancia en la Tierra antes de desaparecer. Los intentos más serios de resolver el misterio se enfocan en las condiciones generales.
No fueron los dinosaurios en sí los que fallaron sino que se trató de algo de mayor importancia. Especies de animales que habían sobrevivido durante millones de años tienen que haberse adaptado bien al medio ambiente, pues de otro modo no hubiesen sobrevivido sin que cambiaran las condiciones de su existencia. Por lo general, los dinosaurios fueron criaturas con gran éxito.
Debían continuar sobreviviendo en tanto que no cambiaran las condiciones de su existencia. Si se modificaban lentamente, sus organismos podrían no tener tiempo de adaptarse a esos cambios, mediante el normal proceso de la evolución. La extinción se produce a causa de cambios, de cambios repentinos, un tipo de cambios cuyos efectos no pueden ser asimilados por los organismos individuales en el transcurso de pocas generaciones.
Cuando Walter Álvarez encontró la pista, estaba buscando otra cosa. Fue uno de esos descubrimientos casuales que salpican los anales de la ciencia. A partir de 1973, Álvarez y algunos otros colegas acudían cada verano a los Apeninos, en la región de Umbría en el centro del norte de Italia, para estudiar las extraordinarias formaciones de piedra caliza que allí se alzan.
La roca está formada por sedimentos de las profundidades del mar acumulados durante un período de ciento cincuenta y cinco millones de años, desde el jurásico hasta el oligoceno, hace treinta millones de años.
El mar cedió paso a la tierra firme y hace unos pocos millones de años la roca que se formó en el fondo marino fue alzada en un fenómeno de elevación de montañas para quedar expuesta a la erosión durante millones de años. La naturaleza muestra allí una de sus más bellas creaciones, profundos lechos de piedra caliza rosada que brindaban placer estético mucho antes de que llegara la revelación científica.
En la garganta Bottaccione, cerca de la ciudad medieval de Gubbio, esa roca caliza destaca de modo particular y es fácilmente accesible. Y fue allí donde Álvarez aplicó su habilidad como geólogo para estudiar una cuestión relacionada con la historia de la Tierra. Los dinosaurios estaban entonces muy lejos de su pensamiento.
Su interés era paleomagnético. Álvarez y sus colegas estaban buscando pruebas en las rocas que sirvieran como evidencia de una inversión en el campo magnético de la Tierra. Querían comparar las inversiones encontradas en Gubbio con las pruebas de otras inversiones que habían sido detectadas en la corteza de los fondos oceánicos por medio de magnetómetros operados desde los puentes de los buques.
Partiendo de esos descubrimientos confiaba en poder desarrollar, tanto en tierra firme como en el mar, un calendario seguro y completo de las fluctuaciones magnéticas en el curso de la historia de la Tierra. Durante sus investigaciones, los geólogos trataban también de relacionar las inversiones de campos magnéticos con la evolución y la extinción de organismos, como mostraba el registro fósil. La piedra caliza de Gubbio es rica en foraminíferos, criaturas diminutas cuyas conchas de carbonato de calcio son la materia prima de una gran parte de la piedra caliza del mundo.
Con una lupa manual los geólogos pudieron ver la profusión de microorganismos que se abrían camino a través de las capas de piedra arcillosa hasta un punto en el que todos ellos, con excepción de una única especie, desaparecían de modo repentino. Este punto de desaparición de los microorganismos marcaba el límite entre el cretáceo y el terciario, la era de la extinción masiva.
No fueron los dinosaurios en sí los que fallaron sino que se trató de algo de mayor importancia. Especies de animales que habían sobrevivido durante millones de años tienen que haberse adaptado bien al medio ambiente, pues de otro modo no hubiesen sobrevivido sin que cambiaran las condiciones de su existencia. Por lo general, los dinosaurios fueron criaturas con gran éxito.
Debían continuar sobreviviendo en tanto que no cambiaran las condiciones de su existencia. Si se modificaban lentamente, sus organismos podrían no tener tiempo de adaptarse a esos cambios, mediante el normal proceso de la evolución. La extinción se produce a causa de cambios, de cambios repentinos, un tipo de cambios cuyos efectos no pueden ser asimilados por los organismos individuales en el transcurso de pocas generaciones.
Cuando Walter Álvarez encontró la pista, estaba buscando otra cosa. Fue uno de esos descubrimientos casuales que salpican los anales de la ciencia. A partir de 1973, Álvarez y algunos otros colegas acudían cada verano a los Apeninos, en la región de Umbría en el centro del norte de Italia, para estudiar las extraordinarias formaciones de piedra caliza que allí se alzan.
La roca está formada por sedimentos de las profundidades del mar acumulados durante un período de ciento cincuenta y cinco millones de años, desde el jurásico hasta el oligoceno, hace treinta millones de años.
El mar cedió paso a la tierra firme y hace unos pocos millones de años la roca que se formó en el fondo marino fue alzada en un fenómeno de elevación de montañas para quedar expuesta a la erosión durante millones de años. La naturaleza muestra allí una de sus más bellas creaciones, profundos lechos de piedra caliza rosada que brindaban placer estético mucho antes de que llegara la revelación científica.
En la garganta Bottaccione, cerca de la ciudad medieval de Gubbio, esa roca caliza destaca de modo particular y es fácilmente accesible. Y fue allí donde Álvarez aplicó su habilidad como geólogo para estudiar una cuestión relacionada con la historia de la Tierra. Los dinosaurios estaban entonces muy lejos de su pensamiento.
Su interés era paleomagnético. Álvarez y sus colegas estaban buscando pruebas en las rocas que sirvieran como evidencia de una inversión en el campo magnético de la Tierra. Querían comparar las inversiones encontradas en Gubbio con las pruebas de otras inversiones que habían sido detectadas en la corteza de los fondos oceánicos por medio de magnetómetros operados desde los puentes de los buques.
Partiendo de esos descubrimientos confiaba en poder desarrollar, tanto en tierra firme como en el mar, un calendario seguro y completo de las fluctuaciones magnéticas en el curso de la historia de la Tierra. Durante sus investigaciones, los geólogos trataban también de relacionar las inversiones de campos magnéticos con la evolución y la extinción de organismos, como mostraba el registro fósil. La piedra caliza de Gubbio es rica en foraminíferos, criaturas diminutas cuyas conchas de carbonato de calcio son la materia prima de una gran parte de la piedra caliza del mundo.
Con una lupa manual los geólogos pudieron ver la profusión de microorganismos que se abrían camino a través de las capas de piedra arcillosa hasta un punto en el que todos ellos, con excepción de una única especie, desaparecían de modo repentino. Este punto de desaparición de los microorganismos marcaba el límite entre el cretáceo y el terciario, la era de la extinción masiva.
Se comprobó que ninguna inversión del campo magnético coincidía con las devastaciones ocurridas en el período limítrofe entre el cretáceo y el terciario. Pero donde los organismos desaparecían terminaba también la piedra caliza que era cubierta por una capa de tierra arcillosa de color gris rojizo de uno o dos centímetros de grosor. Esa tierra arcillosa estaba casi completamente desprovista de fósiles. Por encima de ella se prolongaba, de nuevo, la piedra caliza, llena de fósiles.
Álvarez se sintió intrigado por la historia que podrían contar aquellas arcillas sobre la extinción de finales del cretáceo. Álvarez, catedrático asociado de Geología en la Universidad de California, en Berkeley, se llevó consigo algunas muestras de aquella arcilla cuando emprendió el regreso a su casa, en 1977, y describió a su padre lo que había encontrado.
Su padre, Luis W. Álvarez era un físico que trabajaba en los Laboratorios Lawrence, en Berkeley, y entre cuyos múltiples honores se incluye un Premio Nobel. Los dos científicos, padre e hijo, se preguntaron asombrados cuántos años podían estar representados por aquella capa arcillosa que separaba dos formaciones de piedra caliza en relación con la vida en la Tierra.
Si se conseguía ese conocimiento se podría llegar también a un entendimiento y comprensión de las extinciones del cretáceo-terciario. Aunque Luis Álvarez no era un geólogo se lanzó de lleno en el problema de la arcilla de Gubbio.
Ése era el tipo de investigación que le iba bien. Se describía a sí mismo como un «cazador de choque», un físico subatómico que buscaba «resonancias». Cazando choques fue como detectó e identificó los rayos cósmicos, lo que constituía su interés principal. La arcilla de Gubbio enfrentó a Álvarez con un nuevo desafío. Lo afrontó desarrollando nuevos métodos de investigación científica. Concibió un método para medir la rapidez con que los sedimentos recogidos en el fondo del mar pasan a formar las capas de arcilla.
Le recordó a su hijo Walter que dado que el polvo cósmico y el material micrometeorítico caen en el planeta a un ritmo más o menos constante, éste debe dejar una marca química con la cual se podría calcular la duración del período de tiempo en los límites del cretáceo. El material químico extraterrestre es químicamente diferente del que se encuentra en la superficie de la Tierra. El escaso elemento iridio, por ejemplo, es varios miles de veces superior en los meteoritos que en la corteza terrestre.
Como el iridio tiene una afinidad por el hierro, como ocurre con otros miembros del grupo de elementos del platino, la mayor parte del iridio original debió de asentarse en el núcleo del planeta conjuntamente con el hierro fundido allí.
Así Álvarez decidió medir la cantidad de iridio contenido en la arcilla. El estudio geoquímico de la arcilla de Gubbio, en el que el equipo de Álvarez estaba buscando iridio o cualquier otra materia extraterrestre reveladora, demostró cantidades tan escasas en la muestra que no podían suponer más de una parte por mil millones.
Pero cuando Álvarez vio su primera «muestra» de iridio se dio cuenta de que el experimento para determinar el período de tiempo límite había fracasado, pues encontró en la arcilla mayor cantidad de iridio del que podría esperarse en base a la caída de polvo cósmico o del normal proceso de la Tierra.
Álvarez se sintió intrigado por la historia que podrían contar aquellas arcillas sobre la extinción de finales del cretáceo. Álvarez, catedrático asociado de Geología en la Universidad de California, en Berkeley, se llevó consigo algunas muestras de aquella arcilla cuando emprendió el regreso a su casa, en 1977, y describió a su padre lo que había encontrado.
Su padre, Luis W. Álvarez era un físico que trabajaba en los Laboratorios Lawrence, en Berkeley, y entre cuyos múltiples honores se incluye un Premio Nobel. Los dos científicos, padre e hijo, se preguntaron asombrados cuántos años podían estar representados por aquella capa arcillosa que separaba dos formaciones de piedra caliza en relación con la vida en la Tierra.
Si se conseguía ese conocimiento se podría llegar también a un entendimiento y comprensión de las extinciones del cretáceo-terciario. Aunque Luis Álvarez no era un geólogo se lanzó de lleno en el problema de la arcilla de Gubbio.
Ése era el tipo de investigación que le iba bien. Se describía a sí mismo como un «cazador de choque», un físico subatómico que buscaba «resonancias». Cazando choques fue como detectó e identificó los rayos cósmicos, lo que constituía su interés principal. La arcilla de Gubbio enfrentó a Álvarez con un nuevo desafío. Lo afrontó desarrollando nuevos métodos de investigación científica. Concibió un método para medir la rapidez con que los sedimentos recogidos en el fondo del mar pasan a formar las capas de arcilla.
Le recordó a su hijo Walter que dado que el polvo cósmico y el material micrometeorítico caen en el planeta a un ritmo más o menos constante, éste debe dejar una marca química con la cual se podría calcular la duración del período de tiempo en los límites del cretáceo. El material químico extraterrestre es químicamente diferente del que se encuentra en la superficie de la Tierra. El escaso elemento iridio, por ejemplo, es varios miles de veces superior en los meteoritos que en la corteza terrestre.
Como el iridio tiene una afinidad por el hierro, como ocurre con otros miembros del grupo de elementos del platino, la mayor parte del iridio original debió de asentarse en el núcleo del planeta conjuntamente con el hierro fundido allí.
Así Álvarez decidió medir la cantidad de iridio contenido en la arcilla. El estudio geoquímico de la arcilla de Gubbio, en el que el equipo de Álvarez estaba buscando iridio o cualquier otra materia extraterrestre reveladora, demostró cantidades tan escasas en la muestra que no podían suponer más de una parte por mil millones.
Pero cuando Álvarez vio su primera «muestra» de iridio se dio cuenta de que el experimento para determinar el período de tiempo límite había fracasado, pues encontró en la arcilla mayor cantidad de iridio del que podría esperarse en base a la caída de polvo cósmico o del normal proceso de la Tierra.

En 1978, Walter Álvarez regresó a Gubbio para hacerse con una colección más completa de muestras de rocas buscada a 350 metros por debajo de la línea del límite del período.
La concentración de iridio que había en ellas se vio que era notablemente constante mientras se iba subiendo en dirección a la franja límite, pero una vez allí aumentaba fuertemente.
La capa de arcilla era treinta veces más rica en iridio que los sedimentos que se hallaban directamente por debajo o por encima de ella.
Cuando los dos Álvarez, padre e hijo, vieron los resultados comenzaron a interrogarse sobre cuál podría ser su significado. No podían imaginarse que la existencia de una cantidad anómala de iridio fuera una coincidencia, precisamente allí, en la capa correspondiente al límite entre el cretáceo y el terciario, precisamente en el tiempo de la extinción masiva. No podían pensar en ningún proceso de depositación normal como factible explicación de lo que estaban viendo.
Estaban seguros de que el iridio en la arcilla de Gubbio tenía que ser un jalón de significativa importancia en la historia de la Tierra. Walter Álvarez hizo su primer comunicado sobre sus descubrimientos en Gubbio en 1979, en una reunión de la Unión Geofísica Americana, en Washington.
Dijo que las pruebas del iridio apoyaban las teorías en favor de que había sido un acontecimiento extraterrestre el que desató en la Tierra los cambios que llevaron a aquella extinción masiva. Al principio él y su padre creyeron que el material llegado a la Tierra procedía de una supernova. Cualquiera que fuera la fuente, si se trataba de una fuente extraterrestre, la intensidad del iridio parecía ser a nivel mundial como en realidad ocurría. E
l equipo de los Álvarez examinó muestras conseguidas en Dinamarca y otras de lugares tan alejados como Nueva Zelanda y pudo determinar que las concentraciones de iridio, inusualmente elevadas, estaban presentes siempre en la misma capa geológica con lo que no cabía duda de que se trataba de un fenómeno global.
Finalmente los Álvarez llegaron a la conclusión de que el impacto de un meteorito, de un asteroide grande de al menos diez kilómetros de diámetro ofrecía un «escenario que explicaba la mayor parte, o toda, la evidencia biológica y física» asociada con la capa limítrofe y la extinción masiva.
El asteroide, abriéndose paso por la atmósfera a 100.000 kilómetros por hora, pudo haber causado un cráter de 200 kilómetros de anchura y enviar a la estratosfera una nube de polvo que se extendería rápidamente hasta envolver por completo a la Tierra, sumiéndola en la oscuridad. Un desastre de mucha mayor magnitud tendría que haberse producido como consecuencia del impacto de un asteroide, tal y como lo veía el grupo Álvarez.
No cabía duda de que el impacto tendría que haber provocado violentos terremotos y monumentales inundaciones en las costas. Tenía que haber lanzado al espacio un volumen de escombros sesenta veces mayor que la masa del propio asteroide. El polvo procedente de este fenómeno podría haber cambiado el día en noche durante varios años.
Sin luz solar cesaría la fotosíntesis, lo que causaría la muerte de las plantas y dislocaría la cadena alimentaria desde su propia raíz. Hubiera sido un desastre sin paliativos. La extinción habría sido la suerte de muchas especies que vivían en la Tierra en el momento de producirse la colisión del asteroide.
La concentración de iridio que había en ellas se vio que era notablemente constante mientras se iba subiendo en dirección a la franja límite, pero una vez allí aumentaba fuertemente.
La capa de arcilla era treinta veces más rica en iridio que los sedimentos que se hallaban directamente por debajo o por encima de ella.
Cuando los dos Álvarez, padre e hijo, vieron los resultados comenzaron a interrogarse sobre cuál podría ser su significado. No podían imaginarse que la existencia de una cantidad anómala de iridio fuera una coincidencia, precisamente allí, en la capa correspondiente al límite entre el cretáceo y el terciario, precisamente en el tiempo de la extinción masiva. No podían pensar en ningún proceso de depositación normal como factible explicación de lo que estaban viendo.
Estaban seguros de que el iridio en la arcilla de Gubbio tenía que ser un jalón de significativa importancia en la historia de la Tierra. Walter Álvarez hizo su primer comunicado sobre sus descubrimientos en Gubbio en 1979, en una reunión de la Unión Geofísica Americana, en Washington.
Dijo que las pruebas del iridio apoyaban las teorías en favor de que había sido un acontecimiento extraterrestre el que desató en la Tierra los cambios que llevaron a aquella extinción masiva. Al principio él y su padre creyeron que el material llegado a la Tierra procedía de una supernova. Cualquiera que fuera la fuente, si se trataba de una fuente extraterrestre, la intensidad del iridio parecía ser a nivel mundial como en realidad ocurría. E
l equipo de los Álvarez examinó muestras conseguidas en Dinamarca y otras de lugares tan alejados como Nueva Zelanda y pudo determinar que las concentraciones de iridio, inusualmente elevadas, estaban presentes siempre en la misma capa geológica con lo que no cabía duda de que se trataba de un fenómeno global.
Finalmente los Álvarez llegaron a la conclusión de que el impacto de un meteorito, de un asteroide grande de al menos diez kilómetros de diámetro ofrecía un «escenario que explicaba la mayor parte, o toda, la evidencia biológica y física» asociada con la capa limítrofe y la extinción masiva.
El asteroide, abriéndose paso por la atmósfera a 100.000 kilómetros por hora, pudo haber causado un cráter de 200 kilómetros de anchura y enviar a la estratosfera una nube de polvo que se extendería rápidamente hasta envolver por completo a la Tierra, sumiéndola en la oscuridad. Un desastre de mucha mayor magnitud tendría que haberse producido como consecuencia del impacto de un asteroide, tal y como lo veía el grupo Álvarez.
No cabía duda de que el impacto tendría que haber provocado violentos terremotos y monumentales inundaciones en las costas. Tenía que haber lanzado al espacio un volumen de escombros sesenta veces mayor que la masa del propio asteroide. El polvo procedente de este fenómeno podría haber cambiado el día en noche durante varios años.
Sin luz solar cesaría la fotosíntesis, lo que causaría la muerte de las plantas y dislocaría la cadena alimentaria desde su propia raíz. Hubiera sido un desastre sin paliativos. La extinción habría sido la suerte de muchas especies que vivían en la Tierra en el momento de producirse la colisión del asteroide.
Pero había muchos escépticos y algunos de ellos querían saber cómo era que el impacto del asteroide no había dejado un cráter conocido. Walter Álvarez comentó que nada le gustaría más que encontrar un cráter que pudiera ser el de su asteroide. Durante la década que siguió a la publicación del estudio, la hipótesis de la extinción por el choque de un asteroide continuó siendo tema de debate entre geólogos y paleontólogos.
Uno de las mayores objeciones a esta hipótesis era que no se conocía un cráter cuyas dimensiones correspondieran al tamaño calculado, que debería tener entre 150 y 200 km de diámetro. Si bien no sería imposible que la Tierra hubiera cambiado desde entonces escondiendo una tal deformación, en 1990 se ubicaron indicios en Haití de un tsunami de grandes proporciones que arrastró residuos de iridio. Buscando estudios geológicos realizados desde los años 1960 en adelante se pudo ubicar un cráter en Chicxulub, en la península de Yucatán, con un diámetro de unos 170 km.
Para algunos científicos, un problema de esta teoría es que la lectura de los registros fósiles sugiere que la extinción masiva de hace 65 millones de años duró cerca de diez millones de años, lo que no cuadra bien con que su causa fuera el impacto. Cuando el mesozoico se aproximaba a su fin, unos pocos dinosaurios de pequeño tamaño habían alcanzado una relación cerebro-peso corporal, o cociente de encefalización, que se podría comparar favorablemente con el de los mamíferos primitivos.
Uno de esos dinosaurios era el Stenonychosaurus inequalus, una especie desenterrada por él en 1968, en unas capas geológicas del cretáceo, cerca del Parque Provincial de los Dinosaurios, en Alberta. Partiendo del cráneo y de algunos otros huesos recogidos, determinó que se había tratado de un carnívoro bípedo que debía de pesar unos cuarenta kilos con un metro de altura y que, con la cola extendida, podía medir desde su extremo a la punta del hocico unos dos metros.
El tamaño de la cavidad craneal indicaba que elStenonychosaurus tenía un cociente de encefalización de 0,3, probablemente suficiente para hacer de él el más inteligente de los dinosaurios, con inteligencia varias veces superior a los demás, y posiblemente tan inteligente como los más avanzados de los pequeños mamíferos que fueron sus contemporáneos.
En los seres humanos el cociente de encefalización es de 7,5. También había otros rasgos que hacían de este diminuto dinosaurio un ser excepcional. Al parecer tenía el pulgar libre, un dedo que podía oponerse a las otras dos garras, lo que sin lugar a dudas le daba una especial destreza y capacidad de manipulación. Es posible, también, que tuviera visión estereoscópica. «Parecía tener -dijo Russell- todos los componentes del éxito que posteriormente hemos visto en el desarrollo de los simios».
Uno de las mayores objeciones a esta hipótesis era que no se conocía un cráter cuyas dimensiones correspondieran al tamaño calculado, que debería tener entre 150 y 200 km de diámetro. Si bien no sería imposible que la Tierra hubiera cambiado desde entonces escondiendo una tal deformación, en 1990 se ubicaron indicios en Haití de un tsunami de grandes proporciones que arrastró residuos de iridio. Buscando estudios geológicos realizados desde los años 1960 en adelante se pudo ubicar un cráter en Chicxulub, en la península de Yucatán, con un diámetro de unos 170 km.
Para algunos científicos, un problema de esta teoría es que la lectura de los registros fósiles sugiere que la extinción masiva de hace 65 millones de años duró cerca de diez millones de años, lo que no cuadra bien con que su causa fuera el impacto. Cuando el mesozoico se aproximaba a su fin, unos pocos dinosaurios de pequeño tamaño habían alcanzado una relación cerebro-peso corporal, o cociente de encefalización, que se podría comparar favorablemente con el de los mamíferos primitivos.
Uno de esos dinosaurios era el Stenonychosaurus inequalus, una especie desenterrada por él en 1968, en unas capas geológicas del cretáceo, cerca del Parque Provincial de los Dinosaurios, en Alberta. Partiendo del cráneo y de algunos otros huesos recogidos, determinó que se había tratado de un carnívoro bípedo que debía de pesar unos cuarenta kilos con un metro de altura y que, con la cola extendida, podía medir desde su extremo a la punta del hocico unos dos metros.
El tamaño de la cavidad craneal indicaba que elStenonychosaurus tenía un cociente de encefalización de 0,3, probablemente suficiente para hacer de él el más inteligente de los dinosaurios, con inteligencia varias veces superior a los demás, y posiblemente tan inteligente como los más avanzados de los pequeños mamíferos que fueron sus contemporáneos.
En los seres humanos el cociente de encefalización es de 7,5. También había otros rasgos que hacían de este diminuto dinosaurio un ser excepcional. Al parecer tenía el pulgar libre, un dedo que podía oponerse a las otras dos garras, lo que sin lugar a dudas le daba una especial destreza y capacidad de manipulación. Es posible, también, que tuviera visión estereoscópica. «Parecía tener -dijo Russell- todos los componentes del éxito que posteriormente hemos visto en el desarrollo de los simios».
Con esta idea en mente, Russell se lanzó a formarse un concepto sobre el tipo de animal que hubiera llegado a ser el Stenonychosaurus en nuestros días si él y los demás dinosaurios no se hubieran extinguido hace sesenta y cinco millones de años.
Dio forma a sus ideas en un modelo tridimensional de fibra de vidrio, creado con la ayuda de Ron Sequin, un taxidermista, y que en la actualidad se encuentra en el Museo de Ciencias Naturales en Otawa. Russell llama a este modelo un dinosauroide, una criatura inteligente que podría haber llegado a ser la culminación posible, en el siglo XX, de una evolución ininterrumpida de los dinosaurios, si éstos hubieran sobrevivido.
El modelo de dinosauroide, según Russell, es el producto de un trabajo de suposiciones y una extrapolación «conservadora» de determinadas tendencias evolutivas. Tomando en consideración la tendencia hacia un mayor cerebro en los dinosaurios, que se había manifestado ya antes de su extinción, el dinosauroide hubiera evolucionado ahora hasta un cociente de encefalización del 7,1, que está dentro del límite de los humanos.
Para dejar sitio al mayor cerebro, el cráneo en el modelo fue construido algo mayor que el delStenonychosaurus y el rostro relativamente más pequeño, como se vio ocurría en la evolución de los simios antropoides.
Los ojos eran grandes óvalos, como es característico en los dinosaurios. La postura era erguida. El cuello más corto de lo que era corriente en los dinosaurios, porque de seguir con el cuello largo les hubiera resultado muy difícil soportar el peso de un cráneo grande al final de un cuello horizontal.
La cola, al hacerse innecesaria como contrabalance para el equilibrio, habría desaparecido y por esa razón el dinosauroide no la tenía. Su cráneo es calvo y sus manos tienen tres dedos, ambas cosas características establecidas de los dinosaurios.
Dado que Russell asume, además, que el animal habría evolucionado y dejado de poner huevos para traer al mundo a sus hijos vivos, el dinosauroide tenía ombligo. Russell creía que las crías serían alimentadas con alimentos previamente masticados o regurgitados, como lo son las crías de las aves, y por esa razón los dinosauroides no tenían pezones.
La falta de órganos sexuales externos del dinosauroide es una característica típica de los reptiles. Cualquiera que fuese la fisiología y la conducta social de los dinosaurios en su época, concluía Russell, el dinosauroide habría evolucionado ya hasta convertirse en una criatura fuerte y vigorosa, de sangre caliente, que probablemente viviría formando comunidad con otros compañeros de caza.
Estas criaturas podrían tener voces semejantes al piar de las aves y, posiblemente, algún tipo de lenguaje, aunque no hay modo de saber a qué nivel de encefalización el lenguaje se hace posible o inevitable.
En el modelo de Russell, que como él dice es conservador, el dinosauroide aún no habría alcanzado niveles de inteligencia humana, pero el científico no excluye la posibilidad de que los descendientes del Stenonychosaurus, o cualquier otro de los dinosaurios mejor dotados cerebralmente, pudieran haber evolucionado hasta convertirse en especies más inteligentes, ingeniosas y sofisticadas que el Homo sapiens.
Dio forma a sus ideas en un modelo tridimensional de fibra de vidrio, creado con la ayuda de Ron Sequin, un taxidermista, y que en la actualidad se encuentra en el Museo de Ciencias Naturales en Otawa. Russell llama a este modelo un dinosauroide, una criatura inteligente que podría haber llegado a ser la culminación posible, en el siglo XX, de una evolución ininterrumpida de los dinosaurios, si éstos hubieran sobrevivido.
El modelo de dinosauroide, según Russell, es el producto de un trabajo de suposiciones y una extrapolación «conservadora» de determinadas tendencias evolutivas. Tomando en consideración la tendencia hacia un mayor cerebro en los dinosaurios, que se había manifestado ya antes de su extinción, el dinosauroide hubiera evolucionado ahora hasta un cociente de encefalización del 7,1, que está dentro del límite de los humanos.
Para dejar sitio al mayor cerebro, el cráneo en el modelo fue construido algo mayor que el delStenonychosaurus y el rostro relativamente más pequeño, como se vio ocurría en la evolución de los simios antropoides.
Los ojos eran grandes óvalos, como es característico en los dinosaurios. La postura era erguida. El cuello más corto de lo que era corriente en los dinosaurios, porque de seguir con el cuello largo les hubiera resultado muy difícil soportar el peso de un cráneo grande al final de un cuello horizontal.
La cola, al hacerse innecesaria como contrabalance para el equilibrio, habría desaparecido y por esa razón el dinosauroide no la tenía. Su cráneo es calvo y sus manos tienen tres dedos, ambas cosas características establecidas de los dinosaurios.
Dado que Russell asume, además, que el animal habría evolucionado y dejado de poner huevos para traer al mundo a sus hijos vivos, el dinosauroide tenía ombligo. Russell creía que las crías serían alimentadas con alimentos previamente masticados o regurgitados, como lo son las crías de las aves, y por esa razón los dinosauroides no tenían pezones.
La falta de órganos sexuales externos del dinosauroide es una característica típica de los reptiles. Cualquiera que fuese la fisiología y la conducta social de los dinosaurios en su época, concluía Russell, el dinosauroide habría evolucionado ya hasta convertirse en una criatura fuerte y vigorosa, de sangre caliente, que probablemente viviría formando comunidad con otros compañeros de caza.
Estas criaturas podrían tener voces semejantes al piar de las aves y, posiblemente, algún tipo de lenguaje, aunque no hay modo de saber a qué nivel de encefalización el lenguaje se hace posible o inevitable.
En el modelo de Russell, que como él dice es conservador, el dinosauroide aún no habría alcanzado niveles de inteligencia humana, pero el científico no excluye la posibilidad de que los descendientes del Stenonychosaurus, o cualquier otro de los dinosaurios mejor dotados cerebralmente, pudieran haber evolucionado hasta convertirse en especies más inteligentes, ingeniosas y sofisticadas que el Homo sapiens.
Pero se supone que el Stenonychosaurus nunca tuvo la posibilidad de llevar a cabo su hipotético potencial.
Parece que ni un solo dinosaurio sobrevivió a las extinciones masivas de finales del cretáceo, ya que no se ha encontrado fósil alguno de esas criaturas en capas geológicas más recientes que las del cretáceo.
Los informes ocasionales de que animales con aspecto de dinosaurios han sido vistos en África, nunca fueron confirmados.
Los informes ocasionales de que animales con aspecto de dinosaurios han sido vistos en África, nunca fueron confirmados.
Aunque pudieran ser encontrados algunos animales monstruosos, con un curioso aspecto de dinosaurios, es altamente improbable que los verdaderos dinosaurios hubieran sobrevivido los últimos sesenta y cinco millones de años sin dejar ningún rastro fósil que hubiese sido descubierto hace ya muchos años. Todo ello si excluimos la remota posibilidad de que huyesen (o los llevasen) fuera de la Tierra.
En la actualidad los dinosaurios viven sólo en las mentes de la especie que, sin duda, debe su existencia a la desaparición del dinosaurio.
Si éste no se hubiera extinguido, si su género hubiese continuado dominando la tierra seca, es muy probable que los mamíferos se limitaran a ser pequeñas criaturas de aspecto de roedores, saliendo de noche y escondiéndose durante el día por temor a los poderosos reptiles. O, en el caso de que los dinosaurios hubieran vivido después del cretáceo para haberse extinguido en época más reciente, la Era de los Mamíferos posiblemente estaría aún en su infancia, sin que existieran todavía ni humanos ni protohumanos.
Tenemos, pues, toda la razón al sentimos fascinados por los dinosaurios y, en especial, por su extinción. Las extinciones masivas, como los científicos están comenzando a reconocer, fueron acontecimientos cruciales en el curso de la vida en la Tierra.
Tras la última extinción del cretáceo, debió de existir una enorme desolación por todas partes, silencio y calma donde antes hubo una vida fuerte y vigorosa. Pero, además, una oportunidad para todos aquellos supervivientes que tuvieran la suficiente capacidad de adaptación para aprovecharla.
Las aves disponían del cielo para ellas solas, tras la extinción del pterosaurio. Los peces, los moluscos y otras criaturas del mar ya no tenían que vérselas con los ammonites ni con losmosasaurios.
Los pequeños mamíferos salieron de su baja existencia; ya no quedaban saurios gigantescos, con la excepción de los cocodrilos, y éstos eran incapaces de mantener la hegemonía que durante tanto tiempo fue patrimonio de sus parientes arcosaurianos. Luis Álvarez se dio cuenta, rápidamente, del paralelismo existente entre la hipotética catástrofe que condenó a los dinosaurios y una guerra nuclear.
Los pequeños mamíferos salieron de su baja existencia; ya no quedaban saurios gigantescos, con la excepción de los cocodrilos, y éstos eran incapaces de mantener la hegemonía que durante tanto tiempo fue patrimonio de sus parientes arcosaurianos. Luis Álvarez se dio cuenta, rápidamente, del paralelismo existente entre la hipotética catástrofe que condenó a los dinosaurios y una guerra nuclear.
En 1982 comparó de modo explícito el impacto de su supuesto asteroide con el potencial destructivo de las armas nucleares, una comparación que nos hace dudar de las esperanzas de que se pueda sobrevivir a una guerra atómica. Eso llevó a meditar a otros científicos. Si el impacto de un asteroide puede producir nubes de polvo globales y hacer bajar las temperaturas, ¿cuál sería el efecto de una guerra nuclear sobre el clima de nuestro planeta?

No hay comentarios:
Publicar un comentario